6.3.3.1 - Les gènes homéotiques
Les premières observations ont été réalisées par William Bateson (fig. 6.42) entre la fin du XIXème et le début du XXème siècle.
Parmi ces observations, il constata que différents organismes présentaient des variations par rapport au plan général de l'espèce. A titre d'exemple, il constata chez différents organismes des modifications du plan d'organisation. Chez les plantes peuvent exister des modifications du nombre de pièces florales à l'intérieur d'une même espèce (fig. 6.43).

Chez les Arthropodes, le nombre de segments corporels ou leur nature même peut varier.
Il proposa d'appeler ce processus de changement : homéose[1]. Il définit ce processus comme la transformation d'une partie du corps d'un organisme en une autre partie habituellement présente dans une autre région de l'organisme.
Retrouvé dans plusieurs embranchements, ce processus a été particulièrement analysé chez la Drosophile. Chez cet insecte, il existe de nombreux cas où les segments antérieurs sont transformés. Il parle alors par extension de transformations homéotiques ou de mutations homéotiques.
On trouve par exemple chez un mutant de drosophile une paire de pattes à la place des antennes. Ce mutant, appelé Antennapedia, a une modification de la partie céphalique en segment ayant des caractéristiques thoraciques (fig. 6.44).

Avec l'avènement des analyses génétiques, puis moléculaires, plusieurs gènes ont été caractérisés chez la drosophile. Parmi ceux-ci, on trouve les gènes découverts par Ed Lewis, Christiane Nüsslein-Volhard et Eric Wieschaus (Prix Nobel 1995) encore appelés gènes homéotiques, puisque leur mutation entraînait des modifications segmentaires.
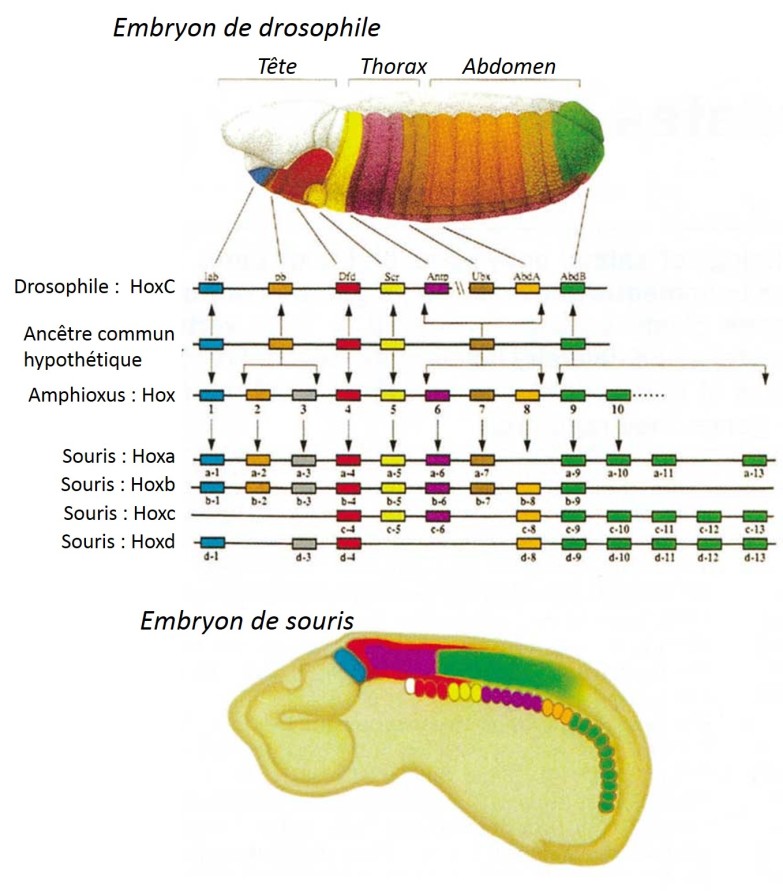
Chez la Drosophile, ce groupe de gènes est situé sur le même chromosome dans un ordre particulier, correspondant à la zone géographique dans laquelle il s'exprime chez l'animal (fig. 6.45). Ainsi, en partant de la partie 3' du locus de ce complexe, on trouve dans l'ordre les gènes responsables de la différenciation antéro-postérieure de l'organisme. Cela s'appelle la colinéarité : la succession de ces gènes est en corrélation avec la région d'expression dans l'organisme ainsi que le moment de leur expression au cours du développement.
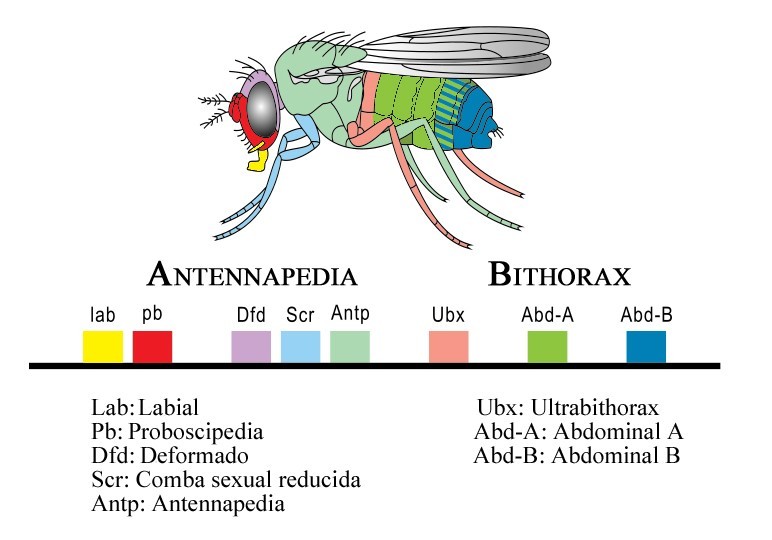
Ces gènes sont au nombre de 8 et organisés en 2 grands ensembles ou complexes : le complexe Antennapedia (ANT-C) et le complexe Bithorax (BX-C). ANT-C est impliqué dans la différenciation et la spécification de la partie proximale, antérieure de l'animal alors que le second complexe est essentiel dans la partie distale de l'organisme depuis la fin du thorax jusqu'à l'ensemble de l'abdomen.
Ces gènes sont importants car ils conditionnent la nature même d'un segment par rapport à l'autre dans l'axe antéro-postérieur et spécifient le type de membres présent sur chacun de ces segments. Étant les premiers gènes découverts responsables de transformations homéotiques, ils ont été dénommés gènes Hom.
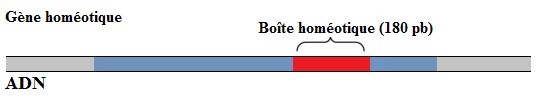
L'analyse fine de ces gènes montre que chacun des gènes du complexe Hom code une protéine capable de réguler la transcription d'autres gènes et surtout que cette action passe par l'existence d'une région de 60 acides aminés capable de se lier à l'ADN appelée homéodomaine (fig. 6.46).
Ed Lewis remarqua que la mutation de certains de ces gènes entraînait des répercussions importantes sur le phénotype. Ainsi la délétion du complexe Bithorax chez la Drosophile conduit à l'apparition d'un embryon avec 10 segments thoraciques identiques ressemblant à un centipède (type mille pattes) (fig. 6.47).

Des atteintes de ces gènes peuvent donc être à l'origine de modifications phénotypiques énormes, similaires à des transformations macro-évolutives.
En étudiant, les autres embranchements, il est apparu que les gènes homéotiques sont présents dans de très nombreux groupes avec des organisations proches de celles retrouvées chez la Drosophile. Ainsi chez les Mammifères comme la Souris, 4 complexes de gènes homéotiques sont retrouvés sur des chromosomes différents. Ils présentent tous comme chez la Drosophile, la séquence codant l'homéodomaine (fig. 6.48).
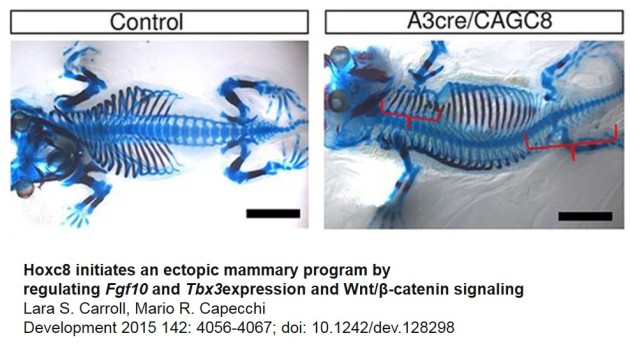
Leurs mutations sont aussi responsables d'altérations développementales selon l'axe antéro-postérieur comme par exemple l'apparition de vertèbres surnuméraires ou la disparition de certaines vertèbres (fig. 6.49).
Ces gènes homéotiques d'organisation similaire à celle de la drosophile mais présents dans d'autres organismes ont donc été nommés par analogie : gènes Hox. Cela renvoie aux notions déjà évoquées au chapitre 3.2.5.2 :
les gènes orthologues qui sont des gènes homologues entre 2 espèces distinctes,
les gènes paralogues qui sont des gènes homologues mais à l'intérieur de la même espèce.
Leur identité de fonction a été montrée par des expériences de sur-expression dans un contexte nucléique différent. Ainsi un gène Hox d'une espèce 1 est introduit dans le génome d'une espèce 2. Dans ce cas, apparaît un segment supplémentaire, ou un organe supplémentaire avec des caractéristiques propres à l'espèce receveuse.
En conclusion, des gènes paralogues d'organisation proche et de fonctions voisines ont été largement répartis chez de nombreux êtres vivants. Ces similitudes impliquent que ces gènes dérivent d'un organisme ancestral ; des analyses plus fines montrent que l'événement initial aurait eu lieu, il y a plus de 500 millions d'années.
Ces gènes de développement intervenant sur des étapes importantes de l'architecture de l'organisme, il est tout à fait envisageable, qu'une mutation sur ces gènes ait des effets macro-évolutifs clairs. Cela peut entre autres expliquer la divergence qui existe entre les Mammifères terrestres et marins au niveau des membres qui ont quasiment disparu, de même que pour les serpents par rapport aux autres Reptiles. Dans ces 2 cas, il pourrait s'agir de mutations dans le complexe Hox 9 à 13.
Limites du concept
Cette hypothèse reste cependant à démontrer et certaines observations la contredisent. Par exemple, les strepsiptères, insectes parasites communs sont caractérisés par une position antérieure des balanciers (fig. 6.50). En quelque sorte, l'identité des segments thoraciques est inversée par rapport à ce qu'on observe chez la drosophile. Pour autant, aucune variation significative de l'expression des gènes Hox n'est corrélé à cette inversion.

La recherche en génétique doit donc encore progresser pour comprendre le fonctionnement et l'origine évolutive des gènes du développement.