Les fractions enrichies en photosystèmes I et photosystèmes II
Comme nous venons de l'expliquer, les chloroplastes contiennent deux types de thylacoïdes à savoir les thylacoïdes non empilés reliant les grana et les thylacoïdes empilés formant les grana (voir le paragraphe "Les différents compartiments du chloroplaste"). Nous avons également vu que ces membranes contiennent l'appareil photosynthétique. Si les propriétés d'organisation des thylacoïdes sont différentes, qu'en est-il de leur contenu ? Pour répondre à cette question, isolons les deux types de thylacoïdes pour en étudier le contenu en protéine. Un protocole typique d'isolement des deux types de thylacoïdes est présenté dans la figure 34A. Il utilise des feuilles d'épinard dont les chloroplastes sont extraits dans du tampon. Les membranes photosynthétiques sont recueillies après un choc osmotique suffisamment intense que pour briser les membranes entourant les chloroplastes et réparties en aliquots. Afin de libérer les protéines enchassées dans les thylacoïdes, les lipides des membranes sont solubilisés à l'aide d'un détergent (ex: Triton X-100). Noter que les détergents sont différents et utilisés dans des concentrations très différentes. Les préparations sont ensuite centrifugées. Selon les traitements, c'est soit le surnageant soit le culot de centrifugation qui est conservé pour l'analyse protéique.
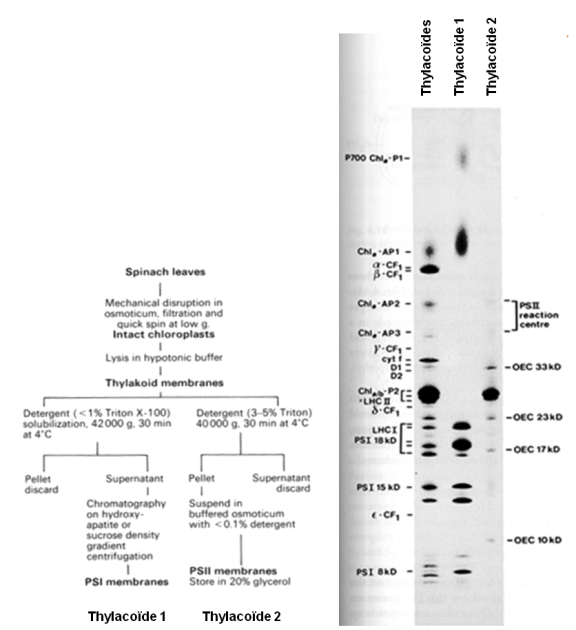
A. Protocole expérimental permettant la préparation des deux types de thylacoïdes. Il est important de noter que la concentration en détergent utilisée pour obtenir les deux types de thylacoïdes est très différente.
B. Comparaison des profils protéiques obtenus avec des fractions contenant les deux types de thylacoïdes (piste de gauche), la fraction “thylacoïde 1” (piste médiane) et la fraction “thylacoïde 2” (piste de droite). Les protéines sont identifiées par leur nom dans les zones bordant les pistes d'électrophorèse. La coloration des bandes de protéines a été réalisée au bleu de Coomassie.
C'est dans la piste de l'extrait contenant les deux types de thylacoïdes que les bandes protéiques sont les plus nombreuses, ce qui est attendu. En revanche, l'extrait “thylacoïdes 1” n'a donné que 9 bandes : le complexe protéique présente la masse la plus élevée correspond aux centres réactionnels des PS I, puis viennent, par ordre décroissant de masse, le complexe apoprotéines-chlorophylles a et les protéines de l'antenne collectrice de l'énergie lumineuse associée au centre réactionnel du photosystème I (Fig. 34B). Dans cette piste, aucune protéine appartenant au photosystème II n'est visible. Cette fraction est donc enrichie en photosystème I.
L'extrait “thylacoïdes 2” présente seulement 5 bandes bien visibles : la bande épaisse regroupe en fait plusieurs protéines de l'antenne collectrice du photosystème II, les autres bandes correspondent à des sous-unités du complexe permettant la production de dioxygène. Dans cette piste, aucune protéine appartenant au photosystème I n'est visible. Cette fraction est donc enrichie en photosystème II (Fig. 34B).
Il est intéressant de noter que certaines protéines présentes dans l'extrait utilisé par la préparation des deux types thylacoïdes ne sont présentes dans aucune des deux autres pistes. Ceci signifie qu'elles ont été perdues lors de la préparation des deux fractions de thylacoïdes. Les protéines concernées appartiennent au complexe cytochrome b6 et f et à l'ATP synthase. Cette expérience démontre qu'en modifiant la concentration en détergent du tampon, il est possible d'isoler, soit les photosystèmes II, soit les photosystème I. Si l'on essaie de comprendre les raisons de ce résultat, le modèle présenté à la figure 35 suivant est obtenu. Il présente la distribution des photosystèmes I et II dans les membranes thylacoïdales. Nous avons constaté que lorsqu'une quantité faible de détergent est ajoutée à la préparation de thylacoïdes, une fraction enrichie en protéines typiques du photosystème I est obtenue. Cela signifie que les thylacoïdes dans lesquels ce photosystème se trouve sont aisément accessibles aux molécules de détergent. Les photosystèmes I sont donc localisés dans les zones exposées au stroma du chloroplaste c'est à dire les membranes extérieures des empilements granaires et les thylacoïdes intergranaires. Au contraire, les protéines typiques du photosystème II ne sont obtenues que lorsque la concentration en détergent est élevée, indiquant que ces protéines sont localisées dans des endroits peu accessibles des thylacoïdes comme le coeur des empilements granaires. Dans le modèle, les photosystèmes II sont notés PSIIα. Ce modèle montre également que les complexes cytochrome b6f sont localisés dans les deux types de membranes alors que ATP synthase (noté dans la Fig. 34B comme CF0-CF1 complex) accompagne les photosystème I. La figure présente également la localisation des PSIIβ qui sont des photosystèmes II n'émettant pas d'oxygène et des antennes collectrices de l'énergie lumineuse détachées des centres réactionnels (notées LHC2 mobile dans la Figure 35). Ces entités apparaissent en cas de stress. Nous en reparlerons à la fin de ce cours. Il est important de noter que ce modèle n'est valable que chez les plantes terrestres et les algues vertes.
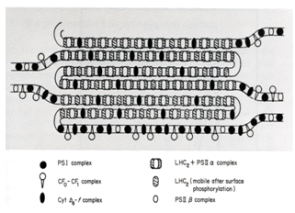
Les photosystèmes I et l'ATP synthase (CF0-CF1 complex) sont localisés dans les zones exposées au stroma du chloroplaste c'est à dire les membranes extérieures des empilements granaires et les thylacoïdes intergranaires tandis que les photosystèmes II sont dans les empilements granaires (PSIIα). Les complexes cytochromes b6f sont localisés dans les deux types de membranes. Les PSIIβ représentent des photosystèmes II n'émettant pas de dioxygène et des antennes collectrices de l'énergie lumineuse détachée des centres réactionnels qui se forment apparaissent en cas de stress.