Le photosystème II
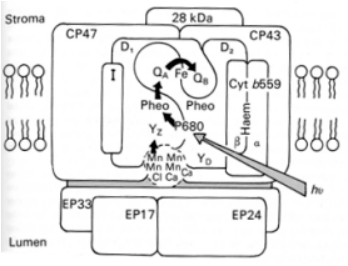
Le photosystème II est composé d'un centre réactionnel, le dimère D1 et D2, et d'une antenne collectrice de l'énergie lumineuse interne, CP43 et CP47, et externe, figurée par le polypeptide de 28 kDa. D'autres polypeptides tels que la protéine I et le cytochrome b559 sont également présents et remplissent des rôles structuraux et/ou de transfert d'électrons autour du photosystème II. Le centre réactionnel et les antennes collectrices sont disposés dans la membrane photosynthétique alors que les protéines impliquées dans la génération des électrons à partir des molécules d'eau sont localisées dans la lumière des thylacoïdes.
Du point de vue fonctionnel, l'énergie absorbée par l'antenne collectrice de l'énergie lumineuse arrive au niveau du P680 (flèche grise) et provoque la formation d'un cation de chlorophylle au niveau d'une des molécules de chlorophylle de ce dernier. L'électron généré est transféré (flèche noire) vers la phéophytine du polypeptide D1. L'électron ne reste que transitoirement à ce niveau et est transféré à QA puis à QB. Le vide électronique créé au niveau de P680 est comblé à l'aide d'un électron provenant de la photolyse de l'eau. Lors du transfert vers le P680, il passe par un résidu histidine (YZ) du polypeptide D1.
hν représente un quantum de lumière.
Hall and Rao (1994) Photosynthesis. 5th edition, Cambridge Univ Press, Fig 8.7, p163
La figure 27 présente un schéma de l'organisation du photosystème II des plantes terrestres. Celui-ci est composé de protéines et de complexes protéines-pigments. Seules les composés les plus importants sont décrits ici. Le centre réactionnel est composé du dimère formé des protéines-pigments D1 et D2. Au coeur du complexe est logée la paire spéciale de molécules de chlorophylle a dont nous avons parlé précédemment. Elle est appelée P680 en référence à la valeur du maximum du spectre d'action de la photosynthèse mesuré par Emerson. D1 et D2 partagent également un atome de fer (Fe) non hémique[1]. D1 et D2 lient chacun une molécule de phéophytine[2] (Pheo dans la figure 27) et un type de quinone : la plastoquinone QB au niveau de D1 et la quinone QA au niveau de D2 (Figure 26). QA ne peut accepter qu'un seul électron à la fois tandis que QB peux en accepter deux.
Les complexes protéines-pigments CP43 et CP47 forment l'antenne collectrice de l'énergie lumineuse. La dénomination CP signifie Chlorophyll Protein et le chiffre indique la masse moléculaire apparente de la protéine lorsqu'elle est séparée par gel d'électrophorèse SDS-PAGE[3]. Le polypeptide de 28 kDa représente l'antenne externe de l'énergie lumineuse. Le photosystème II contient d'autres protéines telles que le polypeptide I et le cytochrome b559 qui remplissent des rôles structuraux et dans le transfert des électrons autour du photosystème II. La description de ces rôles sort du cadre de cette ressource. Comme l'illustre la figure 27, cet assemblage de protéines est disposé dans la membrane photosynthétique. Pour la compréhension du mécanisme, il est important de comprendre l'orientation des composants du photosystème II à savoir que les quinones se trouvent du côté exposé au stroma tandis que le P680 est situé du côté exposé au stroma. Du point de vue fonctionnel, l'énergie absorbée par l'antenne collectrice de l'énergie lumineuse arrive au niveau du P680 (flèche grise dans la figure 27) et provoque l'ionisation d'une des molécules de chlorophylle de ce dernier. P680 est relié électroniquement à la phéophytine de D1. Dans la figure 27, les transferts d'électrons sont représentés par les flèches noires. Le transfert d'électrons vers la phéophytine de D2 n'est jamais observé. L'électron ne reste que transitoirement au niveau de la phéophytine et est transféré à QA puis sur QB. Pour que QB soit saturé en électrons (QB2-), il faut donc que deux électrons aient quitté successivement P680. Ces électrons sont alors injectés dans la chaîne de transporteurs d'électrons, permettant à QB d'accepter de nouveaux électrons. Nous étudierons dans la section "L'éclairement naturel et la mesure de son intensité". les mécanismes sur lesquels repose ce transfert.
Quoiqu'il en soit, le vide électronique créé au niveau de P680 doit être comblé chaque fois qu'un électron est transféré à la phéophytine. Le comblement du vide électronique rend la molécule de chlorophylle à nouveau électriquement neutre et préserve sa stabilité. Si, au lieu de considérer chaque étape du mouvement des électrons de manière relativement statique comme nous venons de le faire, nous considérons le flux d'électrons, c'est à dire la dynamique du système, il devient rapidement évident que le transfert d'électrons à l'intérieur du photosystème II ne peut être performant que s'il existe une source d'électrons abondante et aisément mobilisable pour le comblement du vide électronique se créant régulièrement au niveau du P680. Cette source d'électrons est composée des molécules d'eau présentes dans la lumière des thylacoïdes (Beebo et al. 2013). Les polypeptides - EP33, EP17 et EP24 - impliqués dans le système de photolyse de l'eau dont nous verrons le fonctionnement dans la prochaine section sont localisés dans le stroma des thylacoïdes. Le transfert de l'électron en provenance de l'eau vers le P680, s'effectue via un résidu histidine (YZ) du polypeptide D1.