

5.6.2- Expérience de lignage cellulaire
La principale critique de l'expérience précédente réside dans l'incertitude de l'origine du tissu induit. En effet l'absence de marqueur cellulaire ne permet pas d'attribuer l'origine du mésoderme à l'un plus qu'à l'autre des tissus recombinés. L'origine ectodermique du mésoderme a pu être montrée grâce aux marqueurs de lignage cellulaire. Ce sont des molécules fluorescentes introduites dans les cellules par micromanipulation.
Depuis les années 1980, on utilise des complexes moléculaires réunissant une molécule fluorescente (par exemple la fluorescéine), une molécule de dextran de 10kDa et une molécule de lysine.
L'avantage d'un tel composé réside dans le fait que la fluorescéine rend les cellules fluorescentes. La lysine permet de lier in vivo le composé aux autres protéines du cytoplasme par des liaisons électrostatiques, ainsi que par des liaisons covalentes après fixation pour l'histologie. Le dextran permet de réunir le tout en une macromolécule non métabolisable et non exportable par la cellule.
Ce composé moléculaire est injecté dans l'œuf fécondé, voire au stade deux cellules. Il se répartit dans tout le cytoplasme puis est partagé par toutes les cellules au cours de la période de clivage. L'expérience de Nieuwkoop a donc été reprise de la manière suivante (Fig.113) :
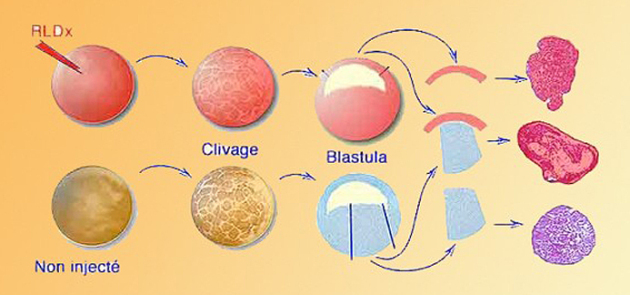
On choisit deux œufs fécondés dont l'un est microinjecté avec le FLDx. Une fois parvenu au stade blastula jeune, on prélève la calotte ectodermique de la blastula marquée (issue de l'œuf microinjecté), et des cellules végétatives de la blastula non injectée. On recombine ces deux explants que l'on met en culture. A l'issue de trois jours de culture, les recombinaisons sont fixées et coupées en histologie. Finalement ces coupes sont examinées au microscope à fluorescence afin de détecter les cellules marquées. Dans ces conditions, on montre que les cellules du mésoderme sont fluorescentes. La conclusion est donc que les cellules mésodermiques sont formées à partir des cellules ectodermiques.
On peut en déduire que, in vivo, l'induction du mésoderme naît des interactions cellulaires entre l'ectoderme et l'endoderme. Ce résultat implique que des réactions similaires doivent avoir lieu in vivo, entre les hémisphères animal et végétatif pendant la période de clivage.
Le principe de l'induction mésodermique est donc le suivant :
L'endoderme est le tissu inducteur. Il porte l'information qu'il transmet à l'ectoderme. On parle de signal inducteur.
L'ectoderme est le tissu inductible. On dit qu'il est compétent, car il est capable de comprendre le signal inducteur, de l'analyser et de répondre par un engagement dans une voie de détermination, en l'occurrence, mésodermique.
L'expérience de Nieuwkoop montre également que la transmission des signaux inductifs nécessite la proximité entre le tissu inducteur et le tissu inductible.
Ceci implique que les signaux inductifs soient des molécules qui diffusent dans l'environnement cellulaire immédiat. On parle d'interactions paracrines.




