

5.6.10 - Rôle des gènes Nodal dans l'induction du mésoderme
Actuellement, ce sont les gènes Nodal de la famille des TGF qui offrent le plus d'évidence quant à leur rôle dans l'induction mésodermique.
Nodal est un gène découvert chez la souris et impliqué dans la formation de l'axe dorsal troncal ainsi que dans les mouvements de gastrulation notamment au niveau dorsal, ce qu'on appelle le « node » équivalent chez le xénope de ce que nous étudierons sous le nom de lèvre dorsale du blastopore.
Xnr1, 2 et 4, dont les ARNm sont maternels et transcrits à la MBT. Ils apparaissent approximativement au stade 9 du clivage et restent présents jusqu'au début de la gastrulation (stade 11) pour Xnr1 et 2, voire jusqu'au stade neurula jeune (stade 14) pour Xnr4. Xnr 2 se maintient à un taux faible jusqu'à la fin de la neurulation. Cependant, on observe que Xnr1 est réexprimé fortement à la fin de la neurulation (stades 17, 18) pour d'autres raisons que celle de l'induction du mésoderme (Fig.123).
On remarquera sur cette image, l'apparition de siamois à partir de la MBT, ainsi que celle d'un gène impliqué dans la formation de la région antérieure, le gène Goosecoïd (gsc) dont nous parlerons un peu plus tard.
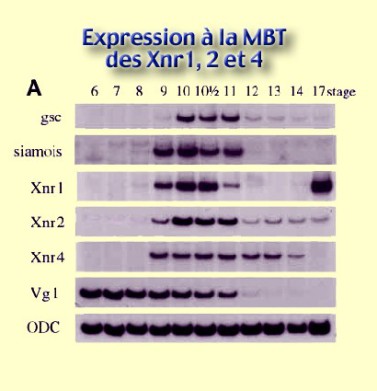
Enfin, on remarquera également que la protéine Vg1 est présente quasiment tout au long de la période de clivage (ici, à partir du stade 6 jeune blastula). L'ODC (Ornithine Décarboxylase) est là pour montrer que les dépôts sont équivalents dans toutes les pistes, contrôle nécessaire pour comparer les pistes entre elles. Ce qui importe pour nous, c'est de pouvoir conclure que les ARNm et les protéines Xnr1, 2 et 4 apparaissent au moment de la MBT.
Régionalisation
L'expression des gènes Xnr1, 2 et 4 s'effectue principalement dans l'hémisphère végétatif et la région dorsale. On le démontre en extrayant les protéines à partir d'explants (Fig.124). Dans une première expérience, on sépare l'ectoderme, la zone marginale et l'endoderme au stade blastula. Xnr1, 2 et 4 s'expriment majoritairement dans l'endoderme et faiblement dans la zone marginale. |  Figure 124 |
Dans une deuxième expérience, on sépare l'endoderme dorsal de l'endoderme ventral. Xnr1, 2 et 4 s'expriment majoritairement dans l'endoderme dorsal. Des traces d'expression sont observées dans la région ventrale, ce qui indique que les Xnr1, 2 et 4 s'expriment faiblement (Fig.125). | 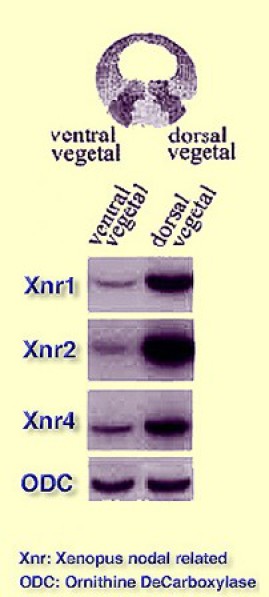 Figure 125 |
En conclusion, Xnr1, 2 et 4 sont exprimés fortement dans l'endoderme dorsal et faiblement dans l'endoderme ventral.
La détection in situ de Xnr1, 2 et 4 montre bien que leur expression est bien endodermique mais située dans la zone marginale. Au début de la gastrulation, elle affecte un gradient dorsoventral décroissant qui apparaît au cours de la période de clivage.
L'expression des gènes Xnr1, 2 et 4 est régulée en amont par VegT. Cette régulation peut être directe on indirecte. On sait cependant, que le promoteur de Xnr1 possède un site de fixation pour le facteur de transcription VegT, ce qui suggère dans ce cas précis que Xnr1 soit directement activé par VegT.
On démontre que les gènes Xnr1, 2 et 4 sont régulés positivement par VegT en utilisant à nouveau les morpholinos. Ainsi, lorsque VegT est déplété, Xnr1, 2 et 4 ne s'expriment plus qu'à un niveau minimum non significatif quelles que soient les concentrations utilisées dans cette expérience (Fig.126). Un argument du rôle de Xnr1, 2 et 4 dans l'induction mésodermique est la restauration des facteurs de croissance par surexpression. | 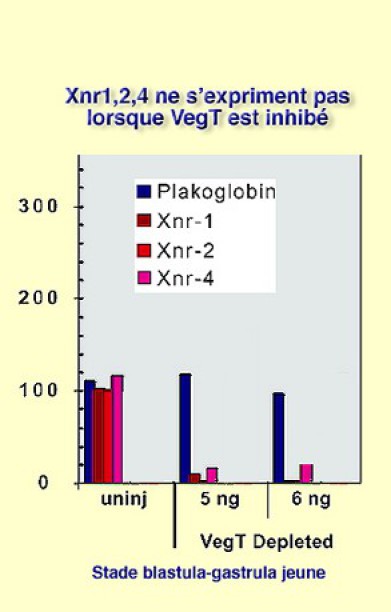 Figure 126 |
Dans un œuf de xénope déplété en VegT on microinjecte soit les ARNm de Xnr1, soit Xnr4, soit l'ARNm de Derrière, un facteur de croissance TGFβ proche de la famille nodal (Fig.127).
A titre ce contrôle, on réalise l'expérience avec les ARNm de eFGF et VegT.
Les deux contrôles positif (uninjected) et négatif (VegT-) donnent le niveau d'expression de 3 gènes sélectionnés pour leur rôle dans la détermination du mésoderme.
Ce sont :
Xbra pour Brachyny, gène ubiquiste du mésoderme qui apparaît à la suite de l'induction du mésoderme. Il peut être considéré comme un marqueurdu mésoderme.
Chordin, induit par l'induction du mésoderme et qui intervient à l'étape suivante dans la dorsalisation du mésoderme et l'induction neurale.
XWnt-8, qui marque le mésoderme latéroventral suite à l'induction mésodermique.
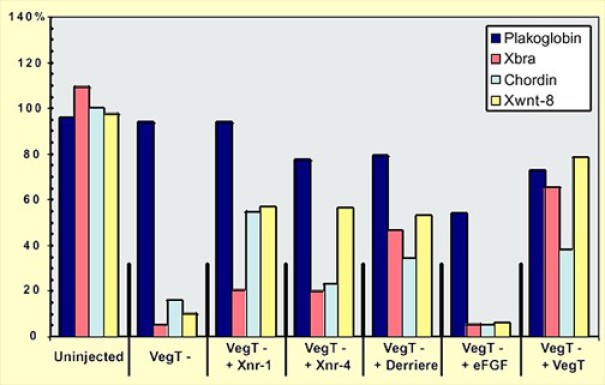
Dans les non injectés, les 3 gènes mésodermique s'expriment au même niveau ou quasiment soit avec un taux aux alentour de 100%.
Dans les injectés avec l'antisens VegT, l'expression de ces 3 gènes tombe à des niveaux proches de 10%.
Si on ajoute Xnr1, Xnr4 et Derrière, on restaure l'expression de ces 3 gènes sans atteindre le niveau des contrôles non injectés (entre 20 et 50 à 60%)
Avec le gène Derrière, on constate que la restauration est plus efficace pour Xbra géne panmésodermique.
Par contre eFGF ne restaure pas l'induction mésodermique médiée par VegT.
Et bien sûr, la restauration avec VegT constitue l'expérience contrôle indispensable.
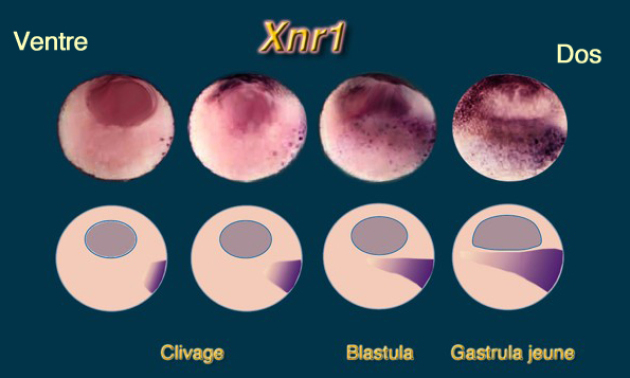
Au niveau morphologique, les facteurs de croissance Xnr1,2,4 s'expriment en un gradient dorsoventral dans l'hémisphère végétatif. Il s'étend au cours de la période de clivage et est réalisé au début de la gastrulation (Fig.128).
En conclusion, cette expérience montre qu'après déplétion de VegT, l'expression des premiers gènes marqueurs du mésoderme restaure l'induction du mésoderme. Par contre, FGf ne fait pas partie de cette voie d'induction et n'est donc pas activé par VegT (Fig.129).
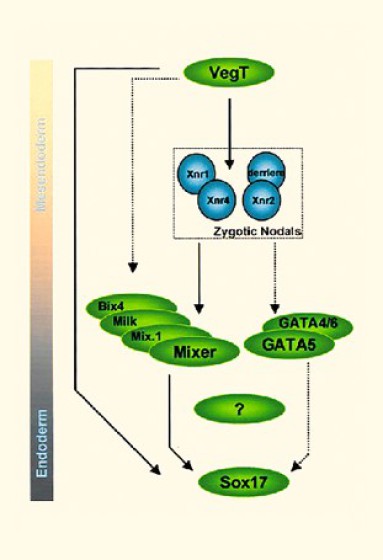
Xnr1 et 4 ainsi que Derrière agissent donc en aval de VegT et sont activés par lui, directement ou indirectement au même titre que les gènes endodermiques Bix4, Mix1 ou Mixer.
Cependant, VegT agit en synergie avec FGF pour l'expression de Xbra.
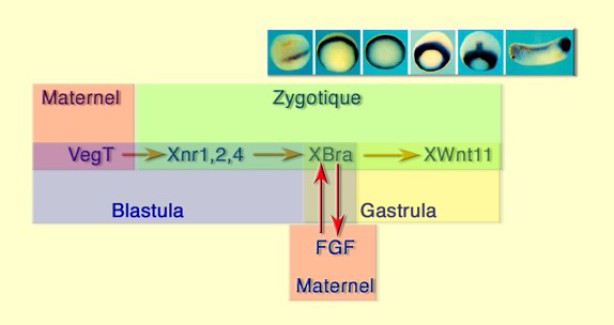
Xbra est un gène ubiquiste panmésodermique. Il apparaît un peu après la MBT au stade blastula 9,5 – 10 dans la zone marginale, se renforce et s'étend dans toute la zone marginale qu'il suit pendant la gastrulation. A la mi-gastrulation il marque la chorde naissante sur le plan médian dorsal puis se restreint à la région caudale qui reste morphogène pendant l'embryogenèse (Fig.130).

Le schéma de régulation de Xbra est le suivant :
VegT active les Xnr1, 2 et 4, lesquels activent Xbra.
Xbra fonctionne alors comme un activateur de FGF qui en retour entretient l'expression de Xbra . nous sommes alors au stade gastrula.
Xbra aura entre autre rôle d'activer l'expression de Xwnt11 dans la zone marginale sachant que XWnt 11 est impliqué dans la polarité et la motilité des cellules du mésoderme, notamment dans les mouvements de convergence extension (Fig.131).
La mutation nulle pour Xbra, ou plus exactement les ARN antisens-Xbra provoquent un phénotype hypomorphe sévère avec absence de région tronco-caudale entr'autres anomalies (Fig.132).
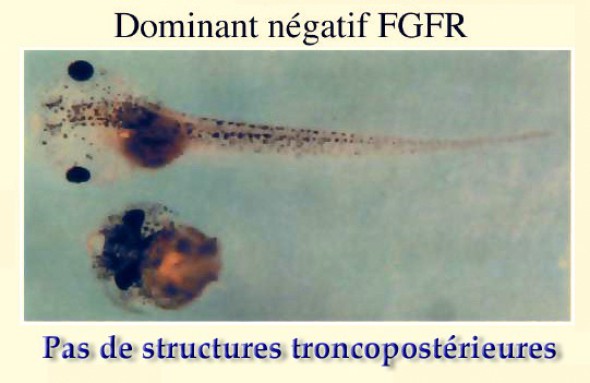
Xbra est donc un des gènes maîtres de la détermination du mésoderme troncal.
En ce qui concerne maintenant Xnr3. Son expression n'est pas modifiée après déplétion en VegT. De plus, la surexpression de Xnr3 ne restaure pas l'induction mésodermique. Par conséquent, Xnr1, 2 et 4 d'un côté et Xnr3 de l'autre ne sont pas régulés par les mêmes voies. En effet, on a vu que Xnr3 et Siamopis sont régulés par les facteurs cat/TCF alors que Xnr1, 2 et 4 le sont par VegT.
Ainsi, Xnr1, 2 et 4 sont impliqués dans l'induction et la régionalisation du mésoderme pendant la période de clivage. Xnr3 apparaît plutôt impliqué dans la dorsalisation et l'induction neurale au cours de la période suivante, la gastrulation.
On peut donc bâtir le modèle suivant (Fig.133) :
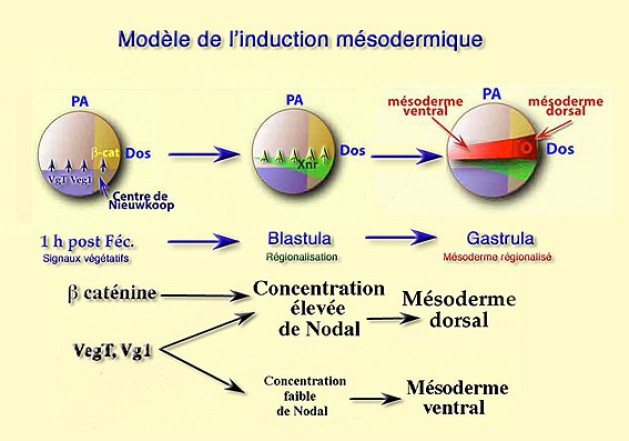
Une heure après la fécondation, la face dorsale est spécifiée par la voie Wut avec la bêta-cat.
Au début du clivage, VegT active les gènes Xnr1, 2 et 4 et leur expression se répartit en un gradient décroissant dorsoventral., dans la région endodermique sous équatoriale, au contact de la zone marginale.
Dorsalement, l'expression élevée de Xnr1, 2 et 4 induisent du mésoderme dorsal.
Ventralement, l'expression faible de Xnr1, 2 et 4 induisent du mésoderme ventrolatéral.
Au stade blastula, la régionalisation a spécifié deux régions mésodermiques dans la zone marginale : le mésoderme dorsal avec un nouveau centre décisionnel : l'organisateur de Spemann et le reste du mésoderme latéroventral non encore déterminé et qui sera séparé en éléments distincts par la dorsalisation au moment de la gastrulation.




