Le déplacement de l'actine induit par la myosine-II
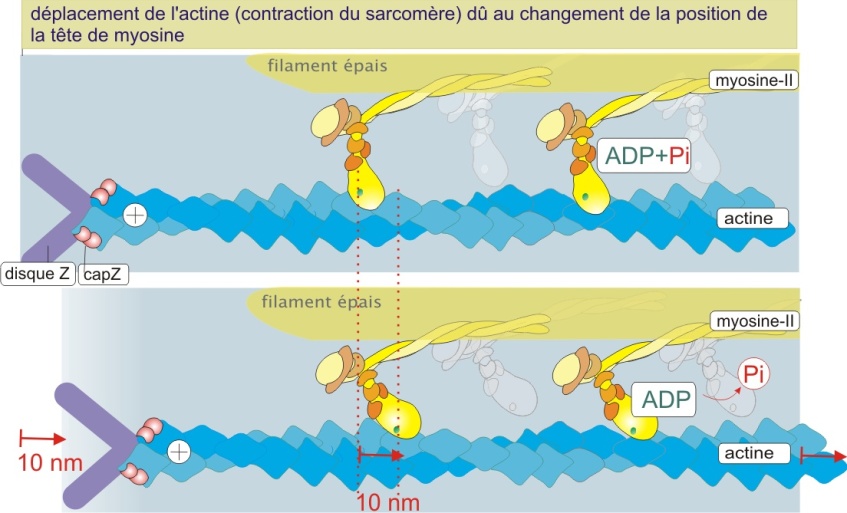
Ce déplacement s'effectue selon un cycle de modifications successives. Au début du cycle, la tête de myosine-II est attachée à l'actine. Cette interaction est de très courte durée car une molécule d'ATP se lie à la tête et provoque une réduction d'affinité pour l'actine. La tête de myosine-II s'éloigne. L'hydrolyse de l'ATP s'ensuit (étape limitante) et induit un changement de la position de la tête de myosine-II (ADP et Pi restent associés à la myosine-II). Dans cet état, la tête s'attache de nouveau à l'actine. La perte subséquente de phosphate (Pi) remet la tête de myosine-II à la position de départ, ce qui déplace le filament d'actine d'environ 10 nm. La perte de Pi est le moment où l'énergie libérée par l'hydrolyse de l'ATP est convertie en mouvement. L'ADP se détache et est remplacé en moins d'une milliseconde par une nouvelle molécule d'ATP et un nouveau cycle peut commencer. La répétition de ce cycle engendre une contraction dynamique (figure 13 ci-dessous).
![]()
Macromedia Flash - 117Ko
![]()
Macromedia Flash - 117Ko
![]() Pour en savoir plus, consultez le document suivant : « Sliding Filaments 50 Huxley [pdf] » (901 Ko).
Pour en savoir plus, consultez le document suivant : « Sliding Filaments 50 Huxley [pdf] » (901 Ko).
Remarque : Du raccourcissement du sarcomère à la contraction du muscle
Le cycle complet se déroule en 50 ms au cours desquelles la myosine-II n'est solidaire de l'actine que pendant 10 ms. Ceci implique qu'une contraction soutenue d'un muscle exige une interaction coordonnée dans le temps et l'espace de plusieurs myocytes. En effet, à tout moment l'interaction actine/myosine-II doit concerner 1/5 du potentiel total. Le sarcomère mesure 3,4\(\mu m\) dans son état étiré et 2,4\(\mu m\) en contraction maximale. En rapide approximation, si on considère que dans un biceps humain de 20 cm les myofibrilles ont 60 000 sarcomères en ligne, on peut dire que ce muscle se raccourcit de 6 cm au maximum. Cette contraction s'effectue en environ 200 millisecondes, en absence de charge. En cas de contraction statique, lorsque le poids de la charge et la force musculaire s'équilibre, l'interaction actine/myosine-II suit le même cycle mais sans déplacement d'actine, c'est-à-dire que la myosine-II doit être alternativement étirée et puis passivement relaxée.
Dans le cas où les stocks d'ATP sont épuisés, la tête de myosine-II reste constamment solidaire de l'actine ce qui résulte en un état de rigidité qui caractérise un cadavre peu de temps après la mort (rigor mortis) !