

6.3.5- Les mouvements de convergence - extension
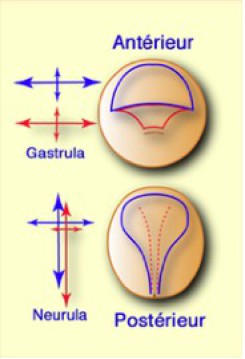
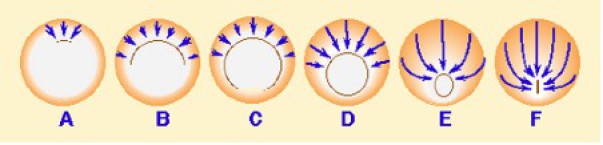
Les expériences de Vogt avaient montré qu'une marque placée latéralement sur la face dorsale effectuait un mouvement de convergence vers le plan médian. Ainsi, les territoires de la chorde et du neuroderme se rétrécissent latéralement. A mesure que le territoire de la chorde pénètre dans l'embryon, celui-ci s'allonge considérablement dans le sens antéropostérieur.
Il en est de même pour le territoire neural qui se rétrécit latéralement, mais reste à la surface dorsale de l'embryon et s'allonge dans la région troncale en prenant une forme dite « en raquette ». L'extension apparaît ainsi comme la résultante de la convergence des tissus vers le plan médian (Fig.171).
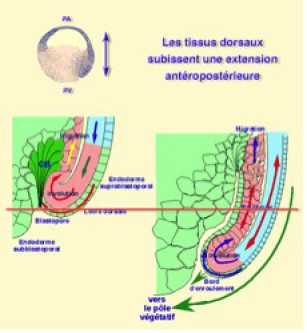
Les tissus dorsaux subissent donc une extension antéropstérieure qui pousse les tissus troncaux vers le blastopore et favorise leur involution sur le bord d'enroulement et de là, leur invagination.
Une fois accrochées à la matrice extracellulaire, les cellules mésodermiques migrent sur le plafond du blastocoele vers le pôle animal (future région céphalique). Ainsi la combinaison des deux mouvements, la convergence extension et la migration antérieure du mésoderme résulte en une extension ou allongement antéroposterieur matérialisé par les doubles flèches rouges du schéma ci-dessous (Fig.172).
Au niveau cellulaire, la convergence résulte de l'intercalation médiolatérale des cellules dorsales aussi bien dans le mésoderme chordal que dans le neuroderme troncal (Fig.173-video5).
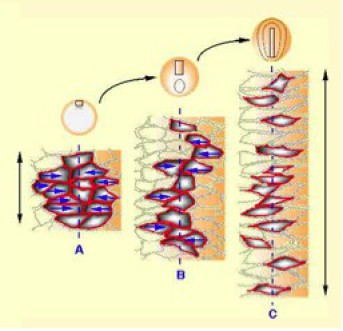
Si l'on considère un explant situé à l'origine, sur la zone marginale dorsale (ZMD), on s'aperçoit que celui-ci s'allonge dans le sens antéropostérieur. Les mouvements de convergence s'expliquent par l'intercalation des cellules dans le plan du tissu (Fig.174). Ils s'effectuent depuis les régions latérodorsales vers la région médiane. On parle d'intercalation médio-latérale.Les mouvements de convergence-extension poussent donc les tissus vers le blastopore.
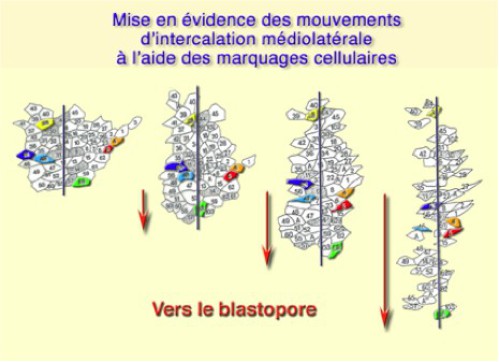
Les tissus soumis à ces mouvements sont amenés à recouvrir le bouchon vitellin et à compléter les mouvements d'étalement de l'ectoderme à la surface de l'embryon, mouvements initiés par l'épibolie (Fig.175-video6).
L'intercalation des cellules entre elles s'explique par des changements d'adhésivité intercellulaire liées à l'expression de molécules d'adhésion cellulaire. Ainsi, deux molécules de la famille des protocadhérines sont impliquées dans ces processus (Fig.176). Chez le xénope, au cours de la gastrulation, l'ensemble du mésoderme dorsal troncal exprime la paraxial protocadhérine (PAPC).
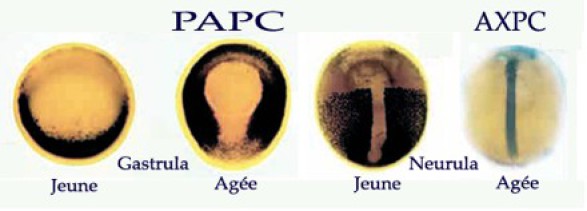
Puis à la fin de la gastrulation, son expression disparaît du territoire chordal mais est maintenue dans le mésoderme paraxial à l'origine des somites.
A cet instant, on voit apparaître l'expression d'une autre protocadhérine, l'axial protocadhérine (AXPC), dans le chordomésoderme. Ces deux molécules s'excluent mutuellement.
L'implication de ces molécules dans les mouvements d'intercalation radiale est attestée par des expériences de surexpression de mutants dominants négatifs (DN-PAPC). Une forme dominante négative de la paraxial protocadhérine exprimée dans les cellules mésodermiques dorsales entraîne l'inhibition de la convergence-extension. Du côté où s'exprime le dominant négatif, les cellules somitiques désorganisées, sétalentt largement latérodorsalement (Fig.177).
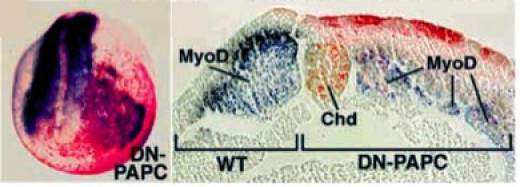
Nous pouvons donc établir une corrélation entre l'expression de molécules liées à la ségrégation d'un territoire à la suite des mouvements de convergence-extension, et la destinée de ce territoire. Ainsi, les cellules de la chorde se séparent de l'ensemble du mésoderme dorsal par un mécanisme de « cell sorting » lié à l'expression de l'AXPC.
L'intercalation médio-latérale s'accompagne d'une bipolarisation des cellules (Fig.178-video7).
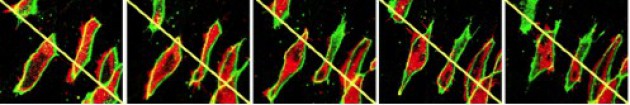
Chaque pôle est animé d'une activité plasmique protrusive intense sous la dépendance d'un facteur de croissance de la famille Wnt, le gène XWnt11.
Wnt11 est exprimé pendant la gastrulation et est activé par Xbra. Il se place donc tout naturellement dans la cascade d'événements initiée par VegT et entretenue par FGF. il faut rappeler que la famille des Wnt se divise en deux catégories (Fig.179).
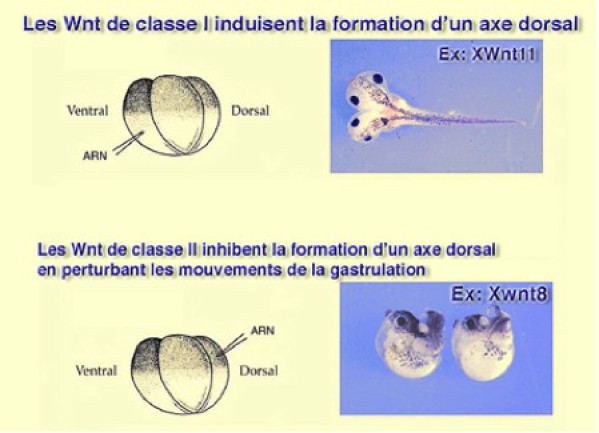
Les Wnt de classe I qui induisent la formation d'un axe dorsal après surexpression
Les Wnt de classe II qui inhibent la formation d'un axe dorsal après surexpression
XWnt11 fait partie de la 1ère catégorie. Quant à la 2ème nous verrons plus tard le rôle de XWnt8. Dans les deux cas rappelons la voie de transduction canonique de la voie Wnt qui aboutit toujours à la libération de la bêta caténine et l'activation d'un gène cible.
On peut corréler XWnt11 à la convergence-extension dans la mesure où le ligand et son récepteur s'expriment dans les tissus dorsaux de la DMZ au moment de la gastrulation (Fig.180).
Ainsi, Xwnt11 est fortement exprimé dans la couche superficielle de la DMZ jusqu'au niveau du blastopore, alors que son récepteur s'exprime dans les cellules profondes du même tissu composé par le territoire de la chorde et du neuroderme (Fig.181).
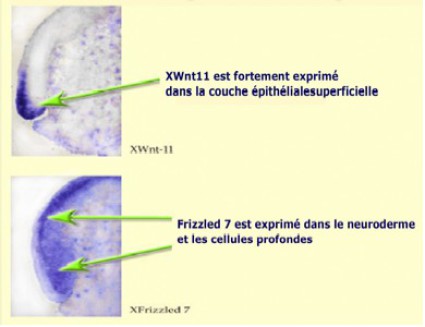
Il semble y avoir une relation topographique étroite entre plusieurs assises cellulaires d'un même tissu, l'une externe et les autres internes qui expriment respectivementle ligand et le récepteur.




