Introduction
Bien que l'anoure Xenopus laevis soit érigé en système modèle pour l'étude du développement embryonnaire des amphibiens (Fig.135), certains mécanismes de la gastrulation sont plus faciles à étudier chez l'urodèle Pleurodeles waltl. Nous nous référerons donc à ces deux espèces.
La gastrulation est caractérisée des mouvements cellulaires coordonnés de garnde ampleur puisqu'ils touchent tous les tissus. La disposition des feuillets est complètement remaniée ce qui génère de nouveaux contacts, donc de nouvelles interactions cellulaires.
Le mésoderme et l'endoderme s'internalisent dans l'embryon. On dit qu'ils s'invaginent. En contrepartie, l'ectoderme s'étale à la surface de l'embryon. On parle de mouvements d'épibolie.
Au total, les trois feuillets originaux s'organisent de manière concentrique :
L'ectoderme limitant externe (à l'origine, par exemple, de la peau)
L'endoderme en position profonde (à l'origine, entre autre, du tube digestif)
Le mésoderme en position intermédiaire (à l'origine, par exemple, du squelette et des muscles).
Une cavité nouvelle est formée dans l'endoderme. Il s'agit du tube digestif primitif auquel on donne le nom d'archentéron.
D'un point de vue mécanistique, les mouvements moteurs de la gastrulation sont principalement assurés par le mésoderme pour les mouvements d'invagination et par l'ectoderme pour les mouvements d'épibolie.
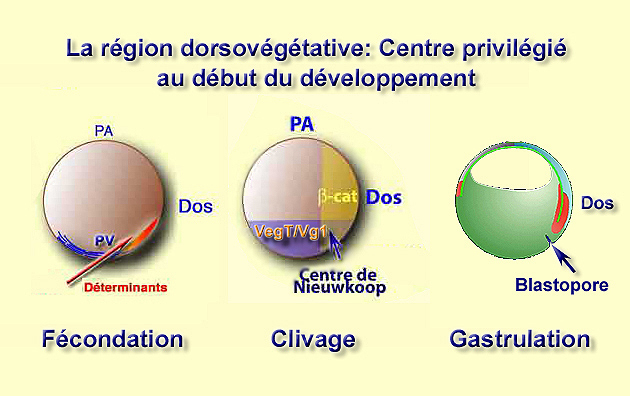
Tout part de la région dorsovégétative que nous suivons depuis la fécondation. En effet, il existe une correlation directe entre la relocalisation dorsoventrale des déterminants végétatifs, la définition du centre de Nieuwkoop et l'apparition du blastopore qui se forme en position dorsovégétative, environ 30° sous le plan de l'équateur (Fig.136).