La fixation du carbone chez les plantes en C4
Nous venons de découvrir que les plantes en C4 présentent la caractéristique de posséder deux types de chloroplastes. A partir des observations de l'ultrastructure de ces deux types de plastes, nous avons fait l'hypothèse qu'aucun des types n'est en mesure de réaliser l'ensemble des réactions photosynthétiques et nous comprenons intuitivement qu'ils vont devoir coopérer. En effet, si les chloroplastes des cellules du mésophylle peuvent réaliser les réactions de la phase photochimique, ils pourraient fournir les produits de ces réactions aux chloroplastes des cellules de la gaine périvasculaire qui en sont incapables mais qui pourraient les utiliser dans le cycle de Calvin-Benson-Bassham. Chez les plantes en C4, nous constatons que les deux phases de la photosynthèse sont délocalisées. Cette conclusion ne permet toutefois pas d'expliquer pourquoi la capacité maximale de fixation du CO2 dans ces plantes est atteinte pour des concentrations faibles en CO2 mais la délocalisation participe à cet effet. Pour expliquer que les plantes en C4 puissent atteindre la capacité maximale de fixation du CO2 pour des concentrations faibles en CO2 il faut imaginer l'existence d'un mécanisme de concentration du carbone. Mettons toutes les pièces du puzzle ensemble et voyons comment cela fonctionne.
Dans les cellules du mésophylle des plantes en C4, le CO2 entre dans le cytoplasme des cellules du mésophylle dans lequel il se dissout. Le CO2 est donc présent sous sa forme hydratée HCO3-. Ce dernier est combiné à une molécule organique, le phosphoenolpyruvate (PEP) (Figure 53) pour former de l'oxaloacétate, une molécule à quatre atomes de carbone. C'est pour cette raison que ce type de photosynthèse est appelé C4. La réaction est catalysée par la PEP carboxylase (Figure 53B). La PEP carboxylase a une grande affinité pour son substrat, l'hydrogénocarbonate et l'incorporation du CO2 peut donc se réaliser même lorsque la concentration en CO2 est faible, par exemple lorsqu'il fait chaud et que les stomates sont fermés. L'oxaloacétate est importé du cytoplasme dans le stroma du chloroplaste dans lequel il est réduit en malate. La réaction utilise du NADPH + H+ formé pendant la phase photochimique de la photosynthèse (Figure 53A). Le malate ainsi formé est transporté des cellules du mésophylle dans les cellules de la gaine périvasculaire. Là, il est décarboxylé en pyruvate qui emprunte le chemin opposé vers les cellules du mésophylle où il sera phosphorylé en PEP, régénérant ainsi le PEP nécessaire à la fixation de nouvelle molécules de HCO3-. Ce cycle est connu sous la dénomination de “cycle de HATCH et SLACK”, du nom des deux scientifiques qui l'ont décrit en 1970 (Hatch et Slack 1970).
Le CO2 issus de la décarboxylation du malate dans le stroma des chloroplastes des cellules de gaine entre dans le cycle de Calvin-Benson-Bassham. Si nous nous rappelons que dans ces cellules, il n'y a pas de production d'oxygène, nous comprenons immédiatement que le risque “photorespiration” est réduit dans ces conditions. De plus, la paroi des cellules de la gaine périvasculaire est imperméable aux gaz, empêchant la diffusion éventuelle du dioxygène ce qui limite encore plus la possibilité de trouver du dioxygène autour de la RuBisCO. L'ensemble de ces faits, en particulier le mécanisme de concentration du CO2 autour de la RuBisCO et l'absence de production d'oxygène dans les chloroplastes réalisant le cycle de Calvin-Benson-Bassham, explique les performances des plantes en C4.
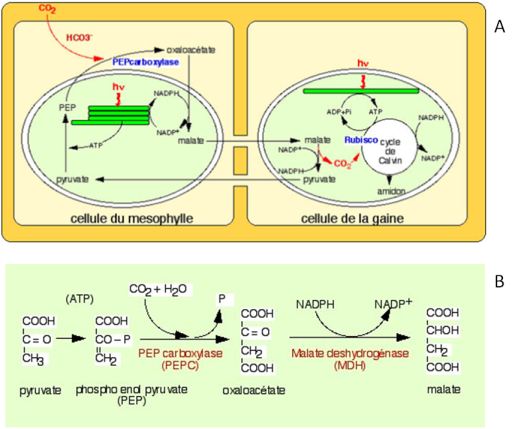
A. Le schéma présente les échanges de composés entre les cellules du mésophylle et les cellules de la gaine. Le CO2 dissout dans le cytoplasme est combiné au phosphoenolpyruvate (PEP) pour former de l'oxaloacétate. Ce dernier est réduit dans le stroma du chloroplaste en malate grâce au NADPH + H+ formé pendant la phase photochimique. Le malate formé est transporté dans les chloroplastes des cellules de la gaine où il est décarboxylé en pyruvate. Le pyruvate est transporté dans les chloroplastes des cellules du mésophylle pour régénérer le stock de PEP. La transformation du pyruvate en PEP nécessite une réaction de phosphorylation pour laquelle de l'ATP est utilisé. Le CO2 issu de la décarboxylation du malate en pyruvate dans le stroma des chloroplastes des cellules de la gaine est utilisé par la RuBisCO, qui le combine avec du RuBP.
B. Schéma des réactions de transformation du pyruvate en malate
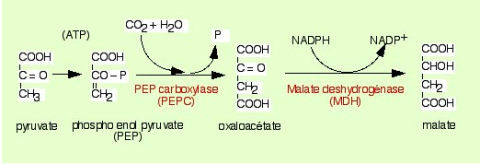
A. Le schéma présente les échanges de composés entre les cellules du mésophylle et les cellules de la gaine. Le CO2 dissout dans le cytoplasme est combiné au phosphoenolpyruvate (PEP) pour former de l'oxaloacétate. Ce dernier est réduit dans le stroma du chloroplaste en malate grâce au NADPH + H+ formé pendant la phase photochimique. Le malate formé est transporté dans les chloroplastes des cellules de la gaine où il est décarboxylé en pyruvate. Le pyruvate est transporté dans les chloroplastes des cellules du mésophylle pour régénérer le stock de PEP. La transformation du pyruvate en PEP nécessite une réaction de phosphorylation pour laquelle de l'ATP est utilisé. Le CO2 issu de la décarboxylation du malate en pyruvate dans le stroma des chloroplastes des cellules de la gaine est utilisé par la RuBisCO, qui le combine avec du RuBP.
B. Schéma des réactions de transformation du pyruvate en malate