Le devenir des trioses phosphates formés dans le cycle de Calvin-Benson-Bassham
Vous vous rappelez que le cycle de Calvin-Benson-Bassham présente une fuite (Figure 44) : à chaque tour du cycle, 1/6 des molécules de triose phosphate quitte le cycle et est engagée dans la production de sucres. Ces sucres peuvent, soit être utilisés immédiatement comme “carburant” dans les mitochondries pour générer de l'ATP et ainsi faire fonctionner le métabolisme, soit être mis en réserve dans le chloroplaste. Le sucre qui est utilisé comme carburant est le saccharose (“sucrose” en anglais) - c'est le sucre que vous mettez dans votre café - et celui qui est mis en réserve est un polymère du glucose appelé amidon (“starch” en anglais) (Figure 49). Vous l'aurez sans doute compris intuitivement, les deux types de sucres ne sont pas synthétisés en même temps ni au même endroit. Le saccharose étant nécessaire lorsque le métabolisme fonctionne, nous ne sommes pas étonnés d'apprendre qu'il constitue la principale destination des trioses phosphates formés pendant que les plantes sont éclairées. Comme précisé dans la figure 49, la transformation des trioses phosphates en saccharose se déroule dans le cytoplasme. Ils doivent donc être exportés du stroma des plastes vers le cytoplasme par le transporteur des trioses phosphates, qui est localisé dans les enveloppes chloroplastiques. Le transport n'est pas gratuit : il se fait uniquement en échange de Pi. Dans le cytoplasme, les trioses phosphates sont pris en charge par la fructose bisphosphatase (FBPase) qui produit du fructose-6-phosphate (F6P). Cette molécule est ensuite transformée en saccharose par la saccharose phosphate synthase (SPS). La réaction produit du Pi, qui sert de monnaie d'échange. La réimportation du Pi dans le chloroplaste est indispensable pour assurer la production d'ATP et la phosphorylation des intermédiaires du cycle de Calvin-Benson-Bassham. Notez que l'activité des enzymes FBPase et SPS est contrôlée de près ou de loin par la présence d'un facteur de l'environnement, la lumière (“diurnal factors” dans la figure 49) et un facteur interne, la disponibilité en fructose-2,6-bisphosphate. Ce composé est un intermédiaire de la glycolyse et gluconéogenèse. Lorsque l'éclairement diminue d'intensité, par exemple en fin de journée, la demande en saccharose est moins importante et il y a moins de Pi produit dans le cytoplasme. Le transporteur des trioses phosphates est donc moins actif. En conséquence, la concentration en triose phosphate augmente dans le stroma du chloroplaste et est utilisé pour la synthèse de l'amidon. Au contraire, la demande en saccharose dans le cytoplasme est importante, par exemple en pleine journée, le Pi est abondamment formé dans le cytoplasme et continuellement échangé contre des trioses phosphates du stroma. Il n'y a que peu de trioses phosphate disponibles pour former de l'amidon. Si pour une raison ou une autre, la consommation du saccharose faiblit dans le cytoplasme, une accumulation transitoire de fructose-1,6-bisphosphate se produit. En conséquence, la quantité de Pi formé dans le cytoplasme diminue et les trioses phosphate s'accumulent dans le stroma du chloroplaste, conduisant à la formation de l'amidon.
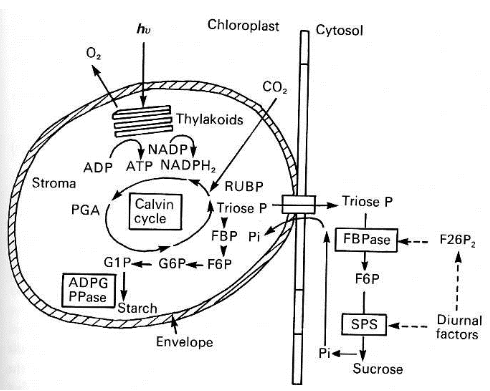
A chaque tour du cycle de Calvin-Benson-Bassham 1/6 des molécules de trioses phosphates quitte le cycle pour satisfaire la demande en saccharose dans le cytoplasme ou/et celle en amidon (“starch” en anglais) dans le chloroplaste. L'exportation des trioses phosphates vers le cytoplasme est assurée par le transporteur des trioses phosphates de l'enveloppe chloroplastique en échange du phosphate inorganique (Pi). Dans le cytoplasme, les trioses phosphates sont transformés en saccharose par la fructose bisphosphatase (FBPase) et la saccharose phosphate synthase (SPS). La réaction produit du Pi. L'activité des enzymes FBPase et SPS est contrôlée de près ou de loin par la présence d'un facteur de l'environnement, la lumière (“diurnal factors”) et un facteur interne, le fructose-2,6-bisphosphate, un intermédiaire de la glycolyse et gluconéogenèse. Comme l'activité de SPS produit du Pi qui est échangé contre les trioses phosphates, l'activation de cette enzyme par la lumière contribue au contrôle de l'orientation des trioses produits dans le chloroplaste vers le cytoplasme. Au contraire, son inactivation bloque l'échange et les trioses phosphates sont utilisés pour la production d'amidon dans le chloroplaste.