4.2.4 - Les séquences ADN confirment l'analyse neutraliste
Le séquençage de l'ADN a permis d'accéder à une somme importante d'informations et a constitué un accélérateur extraordinaire de la compréhension du monde vivant et du développement des biotechnologies. Il a également apporté ou confirmé de nombreuses informations sur le processus évolutif.
La différence majeure entre l'analyse des séquences protéiques et les séquences ADN orthologues[1] tient dans la diversité des pressions de sélections sur ces séquences. Il faut distinguer :
les séquences codantes et non codantes,
géniques et non géniques,
introniques et exoniques
fonctionnelles et structurales
etc.
Une séquence exonique aura globalement un comportement évolutif similaire à celui de la protéine correspondante. Un intron, dont la fonction n'est liée qu'à de courtes séquences caractéristiques évoluera plus librement et donc plus rapidement.
Les séquences intergéniques qui n'interviennent dans aucunes fonctions (hors promoteurs, séquences de régulations, MAR etc.) évoluera beaucoup plus rapidement que les séquences géniques.
Exemple :
Différence de vitesse évolutive selon la position dans le codon
La kératine 9 humaine est une kératine de type 1 exprimée principalement dans les régions palmoplantaires. On trouve un gène orthologue chez le chien.
Si l'on compare la partie de ces gènes codant pour le domaine central de la protéine (les autres régions sont très variables), on observe une homologie globale importante de 85% (635 positions conservées sur 750) (fig. 4.19 et 4.20)
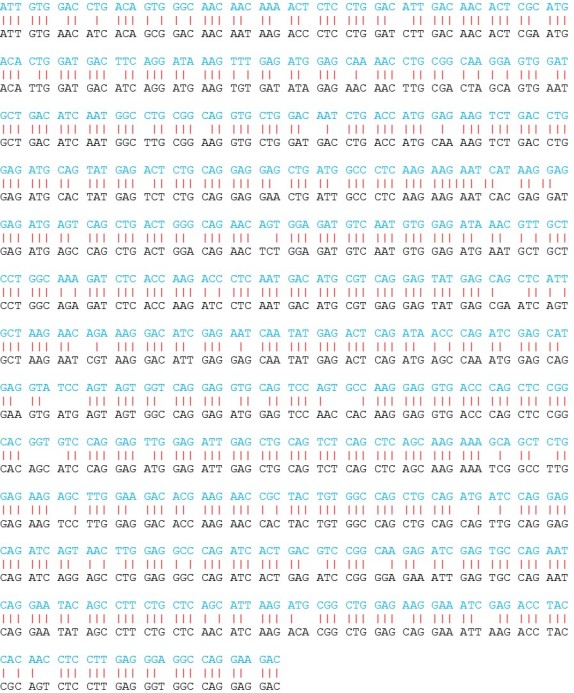
Il est possible d'analyser cet alignement en différenciant les positions correspondant à la première, la deuxième ou la troisième place des codons (fig. 4.20).
taille de la séquence | nb de différences | % des différences | |
|---|---|---|---|
toutes positions | 750 | 115 | 100% |
position 1 | 250 | 36 | 31% |
position 2 | 250 | 25 | 22% |
position 3 | 250 | 54 | 47% |
figure 4.20 : tableau comparatif du nombre de changement dans une partie codante du gène krt9 en fonction de la position sur le codon.
On remarque alors qu'il existe un très net déséquilibre dans la répartition de ces différences :
pratiquement la moitié concernent la troisième position du codon (54/115)
moins d'un quart sont situées sur la première position du codon (25/ 115)
Interprétation
Il convient de mettre ces résultats en rapport avec la configuration du code génétique. Il existe 64 codons dont 61 désignent un acide aminé. Chaque codon peut être muté sur 3 positions (1, 2 ou 3), chaque base peut être intervertie avec l'une des 3 autres (e.g. A peut être remplacé par C, G ou T). Il existe donc sur une séquence codante 549 (61 x 3 x 3) mutations différentes possibles, 183 par position. Du fait de la redondance du code génétique, certaines mutations ne se traduiront pas par un changement au niveau de la séquence protéique (mutation silencieuse). Le tableau ci-dessous (fig4.21) dénombre ces mutations selon la position du codon à laquelle elles se produisent.
type de mutation | silencieuse | faux sens | stop | |||
|---|---|---|---|---|---|---|
position | nombre | % | nombre | % | nombre | % |
| 134 | 24,4% | 392 | 71,4% | 23 | 4,2% |
| 8 | 4,4% | 166 | 90,7% | 9 | 4,9% |
| 0 | 0% | 176 | 96,2% | 7 | 3,8% |
| 126 | 68,9% | 50 | 27,3% | 7 | 3,8% |
figure 4.21 : nature des changement en fonction de la position sur le codon.
La totalité des mutations touchant la position 2 et la très grande majorité de celles touchant la position 1 (95,6%) entraînent un changement d'acide aminé ou l'apparition d'un codon stop.
A l'inverse, plus des deux tiers (68,9%) des mutations de la troisième position n'entraîneront aucun changement de la séquence protéique (mutations silencieuses).
La pression de sélection s'exerçant sur le nouvel allèle généré par mutation dépend de la fonction protéique et donc de sa séquence primaire. On peut faire la prédiction que celle-ci sera très forte sur les positions 1 et 2 et beaucoup plus faible sur la position 3.
La théorie neutraliste postulant que c'est cette pression sélective qui régit la vitesse évolutive d'une séquence, on en conclut donc cette vitesse devrait être lente aux positions 1 et 2 et beaucoup plus rapide en position 3.
A la lumière de cette information, on comprend alors pourquoi il existe plus de changements en position 3 dans la comparaison des kératines de chien et d'Homme :
il est beaucoup plus probable qu'une mutation soit tolérable à cette position pour la fonction protéique.
D'ailleurs si l'on ne tient compte que des mutations faux sens conservées aux trois positions, la différence disparaît (fig.4.22).
tous changements | silencieux | faux sens | |
|---|---|---|---|
toutes positions | 115 | 35 | 80 |
position 1 | 36 | 6 | 30 |
position 2 | 25 | 0 | 25 |
position 3 | 54 | 29 | 25 |
figure 4.22 : nombre de changements silencieux ou faux sens en fonction de la position du codon dans la séquence de krt9
l'excès de différences en position 3 vient des mutations silencieuses.
Cet exemple illustre la plus grande complexité de l'analyse des séquences ADN du fait de la diversité des pressions sélectives pouvant s'exercer sur elles selon leur nature.