4.2.5 - L'horloge moléculaire a des exceptions
L'horloge moléculaire est un concept basé sur l'observation et l'analyse et trouve sa justification dans des considérations statistiques. Cependant d'autres observations montrent que cette hypothèse n'est pas toujours vérifiée. Loin de remettre en cause son principe, il est cependant important de comprendre l'origine de ces exceptions.
1 - L'horloge moléculaire est adaptée aux temps longs
Imaginons une séquence dont l'évolution se produit au rythme moyen de 1 changement par million d'année. Sur une période de 1 million d'année, ce changement peut être observé au début (0,1 Ma) ou au dernier moment (1,0Ma). Il n'est pas improbable, puisqu'il s’agit d'une moyenne, qu'aucun changement ne se soit produit durant cette période ou que plusieurs changements aient eu lieu. Il devient alors très difficile d'analyser la vitesse de divergence de deux espèces particulières sur un tel laps de temps. L'horloge moléculaire peut être vérifiée approximativement sur de longues périodes mais inopérante sur de courtes périodes.
2 - La vitesse évolutive peut dépendre des traits d'histoire de vie
La vitesse évolutive est tributaire de deux paramètres :
le taux de mutation résultant lui même d'un équilibre entre dommages à l'ADN et systèmes de réparation
la vitesse de fixation qui dépend principalement de la dérive génétique et du contrôle exercé par la sélection
Si l'un de ces paramètres varie d'une espèce à l'autre, alors la vitesse évolutive devrait varier.
Les facteurs susceptibles d'intervenir sont entre autre :
les conditions environnementales de vie qui pourraient jouer sur :
l'exposition aux agents mutagènes
la nature et la force des pressions sélectives
l'efficacité des systèmes de réparation ou de protection contre les mutagènes
la nature du métabolisme (e .g. production de ROS)
les modalités de reproduction :
nombre de génération cellulaire dans la lignée germinale
durée de chaque génération
Exemple : influence du temps de génération
Rappelons-nous que la dérive génétique est la variation aléatoire d'un caractère d'une génération à l'autre. Elle dépend :
des événements se produisant durant la vie des individus
de la transmission des allèles entre génération
Sur un siècle, il est possible d'avoir environ :
600 générations de souris
4 à 5 générations d'humains
Les possibilités de dérive sont donc bien plus importantes chez la souris que chez l'Homme :
→ Les séquences de souris devraient évoluer plus rapidement que les séquences humaines.
CONCLUSION
Une espèce ayant un temps de génération long devrait donc évoluer plus lentement qu'une espèce à génération courte.
Dans les faits :
Durant l'analyse des séquences protéiques, cette hypothèse n'est pratiquement pas vérifiée. Si l'analyse des séquences géniques montre une tendance compatible avec l'augmentation des vitesses évolutives chez certaines espèces à génération courte (e.g. les rongeurs), l'ampleur est très loin de ce qui pourrait être envisagé.
3 - Le changement de fonction ou la perte de fonction induit un changement de vitesse évolutive
Comme nous l'avons vu au chapitre précédent, la vitesse évolutive d'une séquence dépend grandement de sa fonction. Dans une lignée évolutive, une espèce particulière, cette fonction peut :
changer de nature
se complexifier, interagir avec d'autres fonctions
disparaître
Les pressions de sélections vont alors changer, se durcir ou au contraire se relâcher. La vitesse évolutive dans cette lignée ou espèce s'en trouvera modifiée.
Exemple : Exemple de la synthèse de vitamine C
La vitamine C joue un rôle important dans diverses réactions physiologiques, telles que la synthèse du collagène, le métabolisme du fer, l'immunité etc. Pour autant, les grands singes, le cochon d'Inde et d'autres espèces encore ont perdu la capacité de sa synthèse. Elles doivent dont trouver la vitamine C dans leur nourriture..
Chez l'homme, le chimpanzé et le cochon d'Inde, l'incapacité à synthétiser la vitamine C est liée à une déficience dans la L-glucono-γ-lactone oxydase (GLO), dernière enzyme de sa chaîne de biosynthèse.
la séquence du gène glo a été analysée dans diverses espèces ( Lachapelle & Drouin, 2011[1]).
Sur le principe de parcimonie que vous étudierez dans le chapitre 5, les comparaisons de séquences ont permis de reconstituer les événements qui se sont produits au cours de l'évolution des espèces. Ils sont présentés dans la figure 4.23.
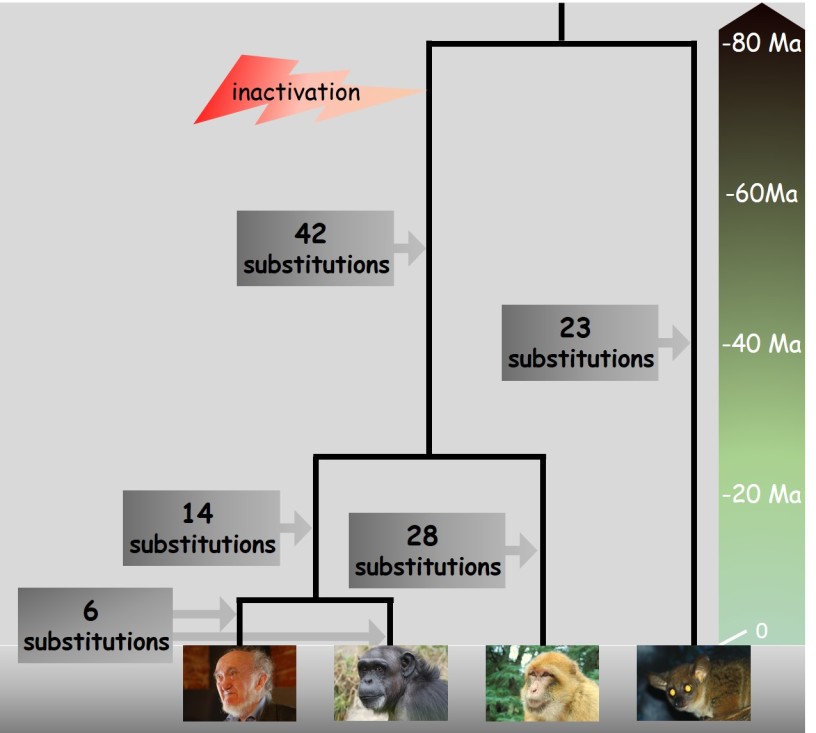
Le gène glo est inactif chez l'Homme, le chimpanzé et le macaque (Catarhiniens) mais parfaitement fonctionnel chez le galago (Otolemur garnettii). Cette inactivation a été datée à moins de 80 Ma, chez un ancêtre commun des trois Catarhiniens.
Il est possible de dénombrer les substitutions s'étant produites depuis la séparation de ce groupe avec la lignée du galago. On compte 61 à 70 positions modifiées chez les Catarhiniens contre seulement 23 chez Otolemur garnettii.
CONCLUSION
La séquence du gène glo évolue environ 3 fois plus vite chez les Catarhinien que chez les galagos. Ce gène étant devenu inutile chez les Catarhiniens suite à la fixation d'une mutation inactivante, la sélection n'a plus jouer son rôle inhibiteur de changement de la séquence, ce qui conduit à une vitesse évolutive supérieure. A l'inverse, le gène glo étant toujours opérationnel chez les primates du nouveau monde, le maintien de la fonction est assurée par la sélection qui conduit à l'élimination des mutations délétères.
On voit donc ici que la perte de fonction d'une séquence modifie drastiquement son comportement évolutif, il en va de même avec toute séquence dont le rôle est plus ou moins modifié dans une lignée particulière.