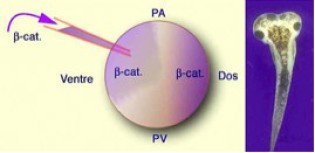
4.4.5-La voie Wnt
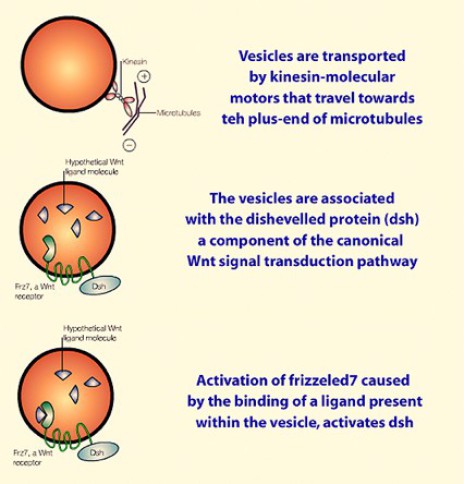
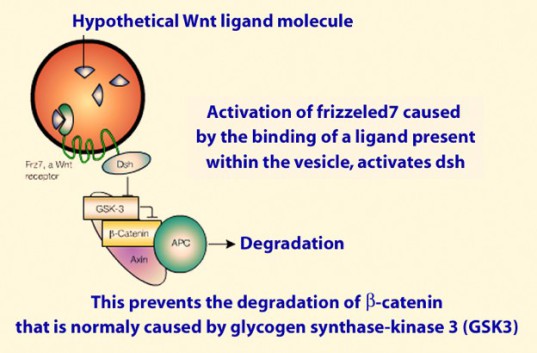
La membrane plasmique de ces vésicules est traversée par des récepteurs transmembranaires de type Frizzled (Frz7) connus pour faire partie de la voie Wnt (prononcer «ouinte»). Or les vésicules golgiennes renferment un Wnt qui se lie à Frz7 et active la voie de transduction Wnt dans le cytoplasme de l'œuf fécondé (Fig.65).
Qu'est ce que la voie Wnt?
Wnt est la combinaison de 2 dénominations Wng (Wingless) chez la drosophile et Int chez la souris.
Les Wnt constituent une grande famille de glycoprotéines, facteurs de croissance qui jouent un rôle dans les développements embryonnaires larvaires et fœtaux, mais aussi dans l'entretien, le renouvellement des tissus adultes. Ils se lient aux récepteurs Fizzled et agissent de façon paracrine pour initier différentes voies de transduction.
Ce que l'on appelle la voie canonique Wnt a pour effet de stabiliser le cofacteur de transcription β-caténine.
Cette voie est la suivante (Fig.66) :
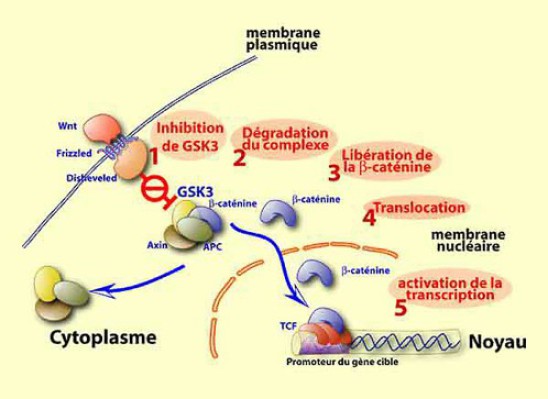
1 - Une fois fixé a son récepteur Frz , le complexe récepteur-ligand active une protéine située dans l'environnement immédiat (c'est-à-dire dans le cytoplasme cortical)= Dishevelled (Dsh).Dsh a pour rôle d'inhiber la fonction GSK3.
2 - GSK3 fait partie d'un complexe de 4 protéines, GSK3, axine, caténine et APC, normalement chargé d'assurer la dégradation de la β-caténine.
Or Dsh activé par Frz envoie GSK3 dans le protéasome pour y être dégradé. Ainsi la β-caténine est stabilisée en tant que facteur de transcription (Fig.67, Fig.68).
3 et 4 - La libération de la β-caténine s'accompagne de sa translocation dans le noyau.
5 - La caténine s'associe à un autre facteur de transcription TCF/LEF pour initier la transcription de gènes cibles consécutifs à la voie Wnt. (TCF : T Cell Factor, LEF : Lymphocyte Enhancer Factor).
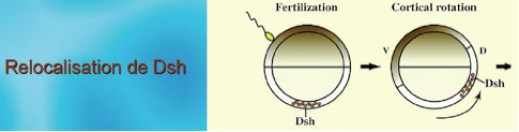
Si l'on replace la voie Wnt dans le contexte de la rotation corticale, on sait que Frz est dans la membrane plasmique des vésicules golgiennes qui renferment un Wnt (vraissemblablement Wnt11). L'activation de Frz par Wnt11 s'effectue au cours de la relocalisation dorsoventrale. Frz active à son tour Dsh dans le cytoplasme cortical.
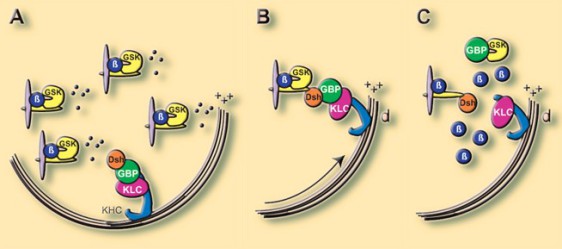
Or, Dsh est couplé à une protéine GBP (pour GSK Binding Protein) elle-même accrochée à la chaîne légère de la Kinésine. (KLC). Et c'est au moment où le complexe formé par Dsh-GBP-KLC-KHC est relocalisé que Dsh est activé par Frz.
A ce moment, Dsh relocalisé, s'associe au complexe GSK3-Axine-APC-Caténine, présente GBP à GSK3 qui l'emmène dans le protéasome. Le complexe ainsi déstabilisé libère la β-Caténine dans la région dorso végétative (Fig.67).
Replaçons ces événements dans l'ensemble de l'œuf fécondé, la cascade des événements est alors la suivante (Fig.69) :
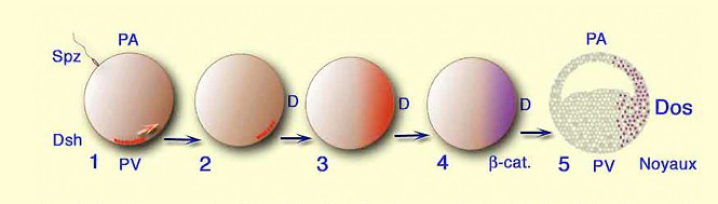 Figure 69 | 1 et 2 - Dsh est relocalisé en position dorso végétative 3 - l'expression de Dsh est amplifiée et s'étend à toute la région dorsale. 4 - La dégradation de de la GSK3 libère la caténine dans toute la région dorsale. 5 - Plus tard, lors de la période de clivage la caténine est transloquée dans les noyaux des cellules dorsales. |
Des arguments en faveur de ce schéma, ont été apportés par les expériences suivantes :
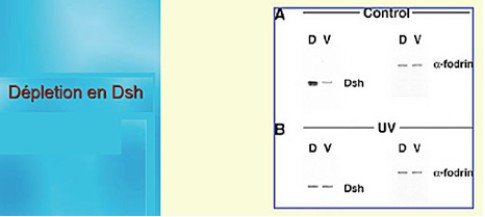
L'irradiation du pôle végétatif provoque une déplétion en Dsh de la face dorsale. En effet, si l'on sépare les moitiés dorsale et ventrale chez les contrôles, on s'aperçoit que seule la moitié dorsale contient Dsh. Par contre, chez les irradiés la moitié dorsale n'exprime pas Dsh et montre un niveau d'expression semblable à celui de la moitié ventrale (Fig.70).
Le couplage de la Dsh et de la GFP (Protéine qui fluoresce en vert) Dsh-GPF permet de suivre le transport de Dsh avec celui des vésicules golgiennes le long des rails de microtubules. Si maintenant, on irradie le pôle végétatif, la fluorescence de la GFP reste au pôle végétatif indiquant que Dsh n'est pas relocalisée en position dorsale.
De même, si on injecte Dsh-GFP dans la région ventrale de l'œuf fécondé, les analyses sur gel de migration permettent de repérer la -caténine stabilisée par Dsh (Fig.72, colonne de gauche). Après irradiation aux UV, Dsh n'est pas activé et la -caténine n'est pas stabilisée et continue d'être dégradée, d'où l'absence de détection sur le gel (Fig.72, colonne de droite).
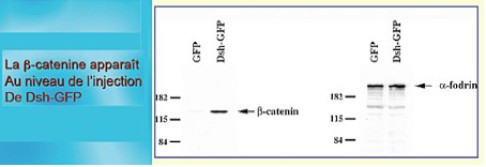
Lorsque l'on injecte de la Dsh-GPF dans la région ventrale (Fig.73, Fig.74)), cette moitié de l'œuf libère la caténine qui n'est pas dégradée. L'injection seule de la GFP donne un résultat négatif, il s'agit de l'expérience contrôle négative.
Lorsque l'on injecte une molécule de la cascade d'événements responsable de l'acquisition de la polarité dorsale comme Dsh ou caténine dans la région ventrale de l'embryon et qu'on laisse évoluer, celui-ci développe du côté injecté (c'est-à-dire ventral) un deuxième axe dorsal antéropostérieur quasi complet (Fig.73).