Une cascade d'évènements
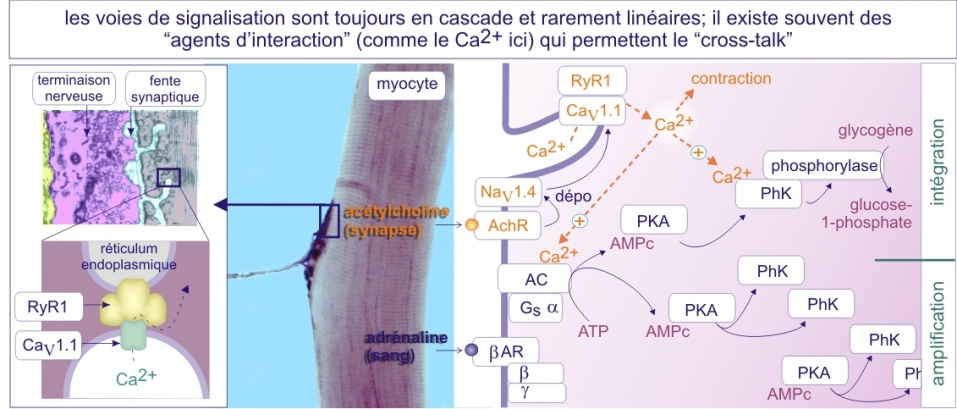
Différents scénarios sont possibles en aval des complexes récepteurs de signalisation. Un cas « simple » concerne des facteurs de transcription (tels que STAT ou SMAD) qui sont directement modifiés en position membranaire, puis transportés vers le noyau où ils changent l'expression de gènes. Dans d'autres cas une longue et souvent étonnante cascade d'événements s'ensuit, avec ou sans production de messagers secondaires, avant que l'enzyme cible ou le noyau soient atteints. De plus, bien que pour des raisons de simplicité nous décrivions les voies de signalisation comme des cascades linéaires, elles constituent en réalité dans la cellule, des réseaux en trois dimensions (« cross-talk »).
Au moins trois raisons peuvent justifier la complexité de telles cascades. D'abord, elles permettraient une rapide amplification; un composant en activant dix, ces dix en activant cent etc. Elles pourraient aussi représenter des sites additionnels de régulation avant que le signal n'arrive sur l'enzyme elle-même (ou entre dans le noyau). Si les cascades étaient strictement linéaires, la compétition pour l'hégémonie ne pourrait se faire qu'au niveau de l'enzyme cible (ou du noyau) ce qui limiterait considérablement les possibilités d'action (voir figure 14). Enfin, les études de modélisation ont montré, plutôt paradoxalement, que les cascades à quatre composants transmettent plus rapidement le signal à l'intérieur de la cellule, que les systèmes à un, deux, trois, cinq ou six composants enchaînés. On peut donc penser que les cellules ont adopté des cascades de trois ou quatre composants uniquement pour réagir rapidement à certains stimuli.
![]() Pour en savoir plus sur le « cross-talk » des voies de signalisation :
Pour en savoir plus sur le « cross-talk » des voies de signalisation :
« Analysis of cross-talk Ranganathan [pdf] » (2,15 Mo).
Complément : Excursion 4 : Sutherland et la découverte du premier « second messager » AMPc.
Tenant compte de l'époque où les connaissances en biochimie des mammifères émanaient souvent d'expérimentations utilisant le foie de rat (perfusion, tranches, homogénats etc), on réalise la chance qui conduisit Earl W. Sutherland à utiliser à la place du foie de rat, du foie de chien pour ses investigations sur la . En effet s'il avait utilisé du foie de rat il n'aurait pas découvert l'AMPc, premier « second messager », car durant son développement le foie du rat perd l'expression des récepteurs \(\beta\)-adrénergiques (qui impliquent la voie de l'AMPc) et la remplace par celle des récepteurs a -adrénergiques (qui impliquent la voie du Ca2+ ).
Bien que les expériences utilisant les hormones (ligands naturels) et les ligands antagonistes radiomarqués, aient permis l'évaluation quantitative des paramètres de liaison aux récepteurs, elles ne permirent pas de les identifier, ni de déterminer leur fonction. Une avance notable est due à Sutherland et ses collègues en 1957 qui montrèrent que la réaction stimulée par l'adrénaline ou le glucagon survient en deux phases. La première réside dans la libération dans le cytoplasme d'un facteur thermostable et dialysable. La seconde, qui se fait uniquement en présence d'ATP, induit la phosphorylation de la phosphorylase b (enzyme qui convertit le glycogène en glucose-1-phosphate). Le facteur thermostable fut identifié comme adénosine monophosphate cyclique (AMPc), le premier « second messager » découvert. Avec cette découverte, il devint possible de relier l'activation de certains récepteurs avec une réponse biochimique spécifique ; un immense nouveau domaine de recherche était alors ouvert (si bien décrit dans le livre « Signal Transduction » par Gomperts, Kramer, Tatham (AP/Elsevier)).
Les travaux d'Earl W Sutherland Jr furent récompensés par l'attribution du prix Nobel de Physiologie ou Médecine en 1971 “for his discoveries concerning the mechanisms of the action of hormones".
Publications historiques :
Rall, TW Sutherland EW, Berthet J. The relationship of epinephrine and glucagon to liver phosphorylase: Effect of epinephrine and glucagon on the reactivation of phosphorylase in liver homogenates. J. Biol. Chem 1957; 224: 463-475.
Sutherland EW. Studies on the mechanism of hormone action. Science 1972; 177: 401-408 .