9.4.2- Les somites
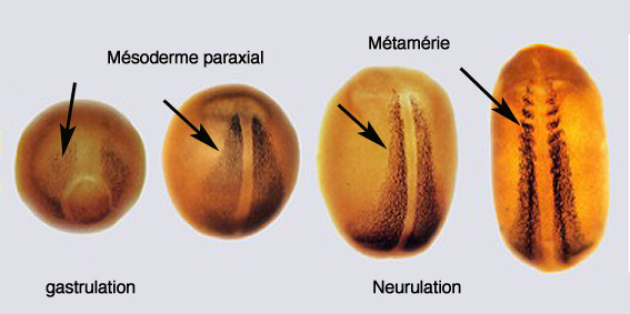
L'émergence des somites est une conséquence de la dorsalisation. Les somites possèdent la particularité de former des unités répétitives le long de l'axe antéropostérieur de part et dautre de la chorde (Fig.270).

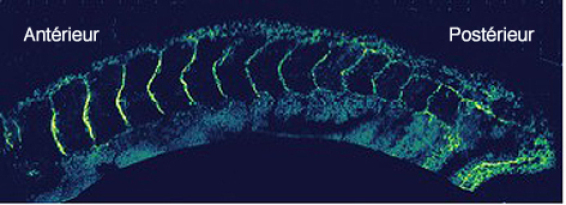
Cette segmentation porte le nom de métamérie. La métamérie est rendue bien visible par la formation des myotomes chez l'embryon d'amphibien (Fig.271). Ceux-ci deviennent perceptibles dès le début du stade du bourgeon caudal.
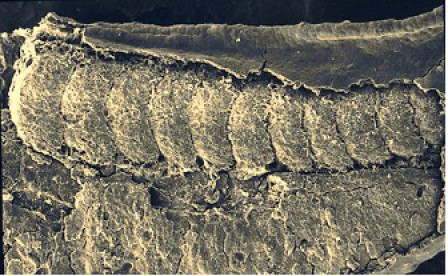
La division de la région antérieure du mésoderme présomitique forme d'abord un somitomère puis la transformation du somitomère en somite est marquée par un changement de polarisation de l'épithélium embryonnaire (Fig.272).
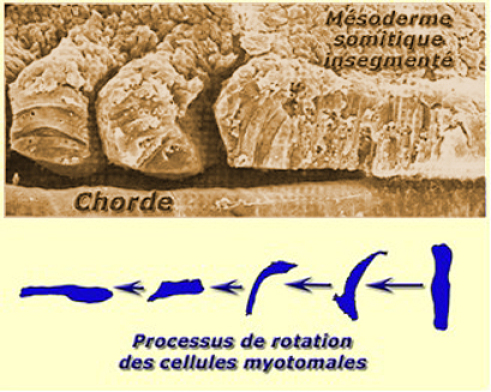
Les cellules présomitiques, primitivement perpendiculaires à l'axe antéropostrérieur, effectuent une rotation de 90°, pour devenir parallèle à l'axe antéropostérieur dans le somite. L'individualisation du somite est concomitante de l'apparition d'une matrice extracellulaire riche en fibronectine qui entoure le somitomère (Fig.273).
Chez les amphibiens comme les autres modèles de vertébrés, la somitogenèse s'accompagne de l'expression des marqueurs de la lignée musculaire.
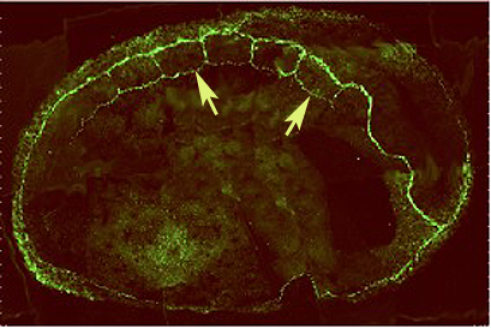
Dès le début de la gastrulation, les zones marginales dorsale et latérodorsale expriment les gènes MyoD (Myoblaste Determinent, Fig.274,A & B) et Myf5 (Myogenic factor 5, Fig.274,C). Au cours de la gastrulation, leurs expressions se restreignent à la zone dorsolatérale, c'est à dire au mésoderme paraxial ou territoire des somites (Fig.275).
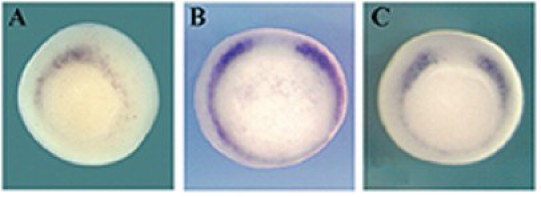
A la fin de la gastrulation et au début de la neurulation le marquage, notamment par MyoD, accompagne la métamérie des somites (Fig.275).
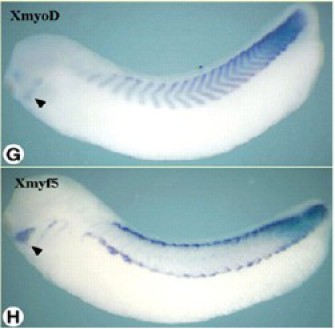
MyoD et Myf5 apparaissent donc comme de bons marqueurs précoces de la somitogenèse. Tout au long de l'organogenèse, ils s'expriment dans les somites et notamment dans la lignée musculaire (Fig.276).
Des études chez le poulet et la souris montrent que MyoD et Myf5 apparaissent liés à la détermination de lignées musculaires différentes, ce qui transparaît sur l'image ci-contre où les marquages respectifs se localisent dans des régions différentes et apparemment complémentaires des somites de Xenopus.
Au sein du mésoderme, la ségrégation du territoire somitique est associée à l'expression de molécules d'adhésion cellulaire proches des cadhérines (Fig.277). Ce sont les protocadhérines. La paraxial protocadhérine (PAPC) s'exprime d'abord dans toute la zone dorsolatérale au cours de la gastrulation puis se restreint à la zone marginale latérale, territoire du mésoderme paraxial somitique.
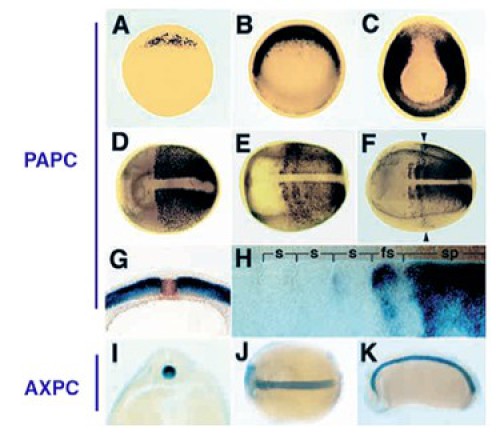
Corrélativement, le territoire chordal se distingue du territoire somitique, en exprimant une autre protocadhérine, l'axial protocadhérine (AXPC). Les protocadhérines sont impliquées dans les mouvements de convergence extension lors de la gastrulation. Des expériences de surexpression d'une forme dominante négative de la PAPC entraîne l'inhibition de l'intercalation médiolatérale.
Ainsi, à la fin de la gastrulation, ces deux territoires sont ségrégés par l'expression différentielle de deux molécules d'adhérence cellulaire. Ces dernières confèrent à chaque tissu un changement d'adhésivité qui conditionne la reconnaissance cellulaire.
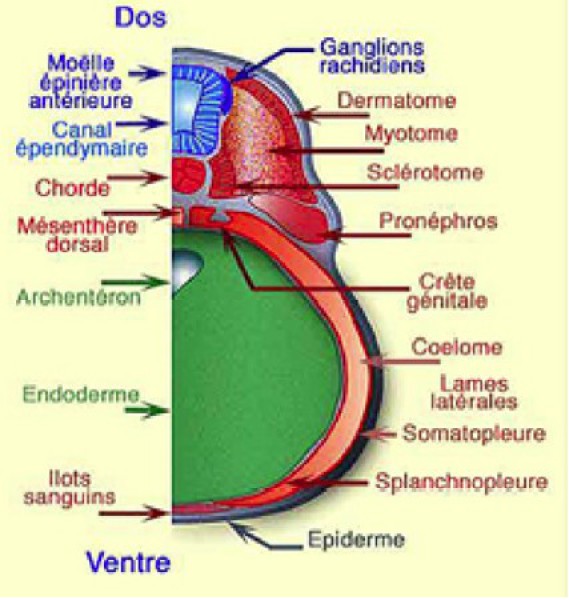
Le somite est une ébauche d'organe embryonnaire transitoire. Outre la lignée musculaire, il est à l'origine de deux autres dérivés: le sclérotome, en position profonde, à l'origine du corps vertébral et le dermatome, superficiel, à l'origine du derme troncal. Le myotome se situe en position intermédiaire (Fig.278).
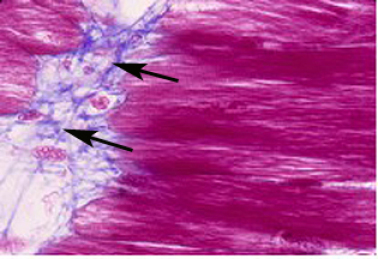
Les myotomes sont constitués de myoblastes qui fusionnent en myotubes syncitiaux pour former la fibre musculaire stiée, polynuclée. Les fibres musculaires, parallèles à l'axe antéropostérieur, sont regroupées en chevrons métamériques (Fig.279). Les espaces extracellulaires entre chaque chevron sont riches en molécules de substrat telles que la fibronectine et la tenascine (Fig.280).
On retrouvera d'ailleurs cette dernière en grande proportion dans les extrémités tendineuses (Fig.281).