Orientation du transport nucléaire : rôle de Ran
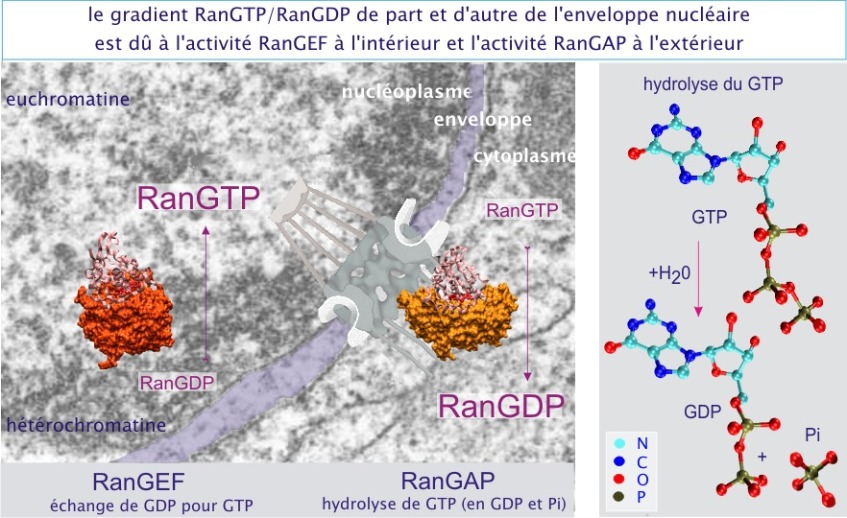
Fondamentalement, le transport nucléaire facilité fait intervenir des récepteurs à double affinité : ils fixent d'une part leur charge (par exemple importine–\(\alpha\)/Rb ou exportine–1/STAT–1) et d'autre part se lient à un motif riche en phénylalanine et glycine (FxFG) localisé sur les protéines du pore. Dans ce processus la GTPase Ran joue un rôle important en déterminant l'affinité du récepteur pour sa charge. Lorsque Ran est fixé au GTP (RanGTP), il entraîne la perte de charge de l'importine–\(\beta\). Inversement, sous la forme RanGDP, il entraîne la perte de charge de l'exportine–1. En conséquence, une fois libérée, la charge ne peut pas revenir en arrière. Ce mode de fonctionnement détermine donc l'orientation du passage à condition que le RanGTP soit abondant dans le nucléoplasme et rare dans le cytoplasme, et inversement pour RanGDP. Ce gradient de RanGTP/RanGDP est mis en place grâce à une forte activité enzymatique qui échange GDP pour GTP dans le nucléoplasme, et une forte activité enzymatique qui hydrolyse le GTP (en GDP + Pi) dans le cytoplasme. L'échange est réalisé par le RanGEF (guanosine exchange factor) et nécessite la présence d'histones (protéines liées à l'ADN pour former la chromatine), donc coté nucléoplasme. L'hydrolyse elle même, est effectuée par le RanGAP (GTPase activating protein) et nécessite la présence d'une protéine filamenteuse cytoplasmique (Nup358 aussi appelée protéine de liaison de Ran ou RanBP) (voir figure 11).
Il faut noter que la famille des importines (adaptateurs et récepteurs) chez l'Homme contient au minimum 17 membres et que chaque membre peut avoir une préférence pour une charge ou une autre. Dans leur ensemble ces protéines sont qualifiées de karyophérines.