4.3.2 - L'évolution est un processus continu : cas des "fossiles vivants"
Le terme a été forgé par Darwin pour désigner des espèces semblant n'avoir pas évolué depuis des millions d'années « pour avoir vécu dans des zones confinées et, de ce fait, avoir été exposées à une compétition moins sévère »
"
Un fossile vivant serait :
Une espèce pour laquelle peu ou pas d'espèces apparentées existent encore
une espèce qui n'a pas ou peu changé morphologiquement depuis un ancêtre lointain
Les critères utilisés sont donc :
l'ancienneté supposée d'apparition du groupe phylétique
la rareté des espèces apparentées, critère subjectif sans fondement scientifique. En effet de nombreuses espèces appartenant à des phyla communs pourraient mériter la même dénomination.
la constance des caractères visibles au cours du temps, critère le plus discutable qui peut être illustré par une exemple célèbre, celui du cœlacanthe.
Attention :
le concept est en totale contradiction avec les faits observés :
l'évolution est un processus continu, l'adaptation par sélection, responsable des changements morphologiques, n'est pas le moteur exclusif. L'analyse des séquences ADN en a fait la démonstration.
même les plus célèbres d'entre eux sont entachés d'inexactitudes sur la ressemblance supposée entre les formes fossiles et actuelles sensées démontrer l'absence d'évolution.
Exemple : Exemple du cœlacanthe : a-t-il si peu évolué ?
Les cœlacanthes sont des Ostéichthyens (poissons osseux) dont les fossiles retrouvés sont nombreux (fig4.25). On croyait cet ordre disparu depuis longtemps.

Cependant, en 1938 des pêcheurs ont remonté dans leurs filets le premier individu vivant connu de l'espèce Latimeria chalumnae (fig. 4.26).

En 1998 une autre espèce du même genre (Latimeria menadoensis) a été à son tour découverte (fig. 4.27).

La relative ressemblance entre la forme moderne et les formes fossiles a conduit à considérer le cœlacanthe comme l'archétype du fossile vivant ou pire comme l'argument réfutant la théorie de l'évolution pour certains créationnistes.
Un article très intéressant dans la revue Bioessays[1] met cependant à mal ce concept, tout particulièrement sur le cas du cœlacanthe.
Certaines études du génome des cœlacanthes modernes ont suggéré une vitesse d'évolution moléculaire lente et un rapport entre cette vitesse et la supposée stase[2] morphologique de ces organismes. Plusieurs points sont discutables dans cette analyse :
la faible vitesse évolutive constatée par rapport aux autres espèces apparentées dépend largement des séquences choisies et ne constitue pas une règle générale
la stabilité morphologique des cœlacanthes n'est qu'un mythe :
aucun fossile ressemblant sérieusement aux espèces modernes n'a été trouvé. Si une analyse grossière permet de constater une parenté entre le fossile présenté figure 4.25 et Latimeria, il est à noter que ce dernier est pratiquement 3 fois plus grand que l'espèce fossile.
les cœlacanthes ne représentent pas un groupe restreint, de nombreux genres morphologiquement différents ont existé
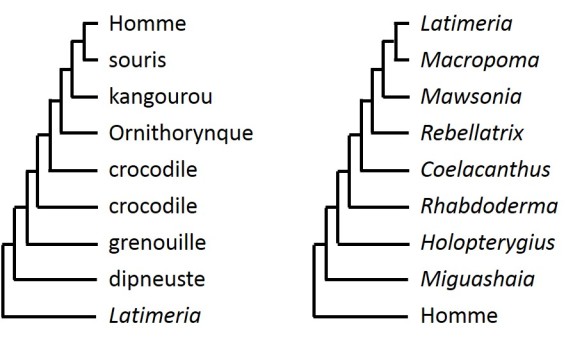
la notion d'"espèce ancêtre" est largement dépendante de la représentation phylogénétique choisie (fig. 4.28) :
si l'on développe la phylogénie de la lignée menant à l'Homme (à gauche dans la figure 4.28), il est aisé de confondre la bifurcation précoce des cœlacanthes avec une "origine ancienne" et le maintien de caractères ancestraux.
si, à l'inverse on choisit de développer la phylogénie des cœlacanthes (à droite dans la figure 4.28), il paraît beaucoup plus clair que le fait qu'une espèce qui se sépare des autres précocement ne représente en rien une forme ancienne de vie.