5.1.3.2.4 - Datation
Jusqu'à l'avènement de la biologie moléculaire et la mise à disposition de nombreuses séquences ADN ou protéiques, seules les données paléobiologiques et les méthodes physiques permettaient une datation des divergences entre espèces.
L'hypothèse de l'horloge moléculaire étudiée dans le chapitre précédent, postule sur une accumulation relativement constante des changements de séquences des macromolécules au cours du temps. Le nombre de différences entre les séquences orthologues de deux espèces peut alors servir de mesure de temps de séparation. La contrainte est de pouvoir calibrer cette horloge. Les données paléontologiques restent donc essentielles.
Méthode :
Une phylogénie de plusieurs espèces (6 dans l'exemple fig. 5.21) peut être établie en comparant des séquences orthologues et en comptabilisant les différences et ressemblances. Un arbre du type de celui-ci dessous peut être établi.
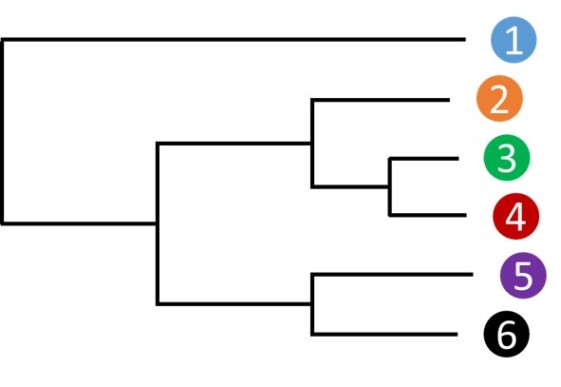
Si l'horloge moléculaire est parfaitement respectée, la distance entre deux espèces et l'un de leurs ancêtres communs doit être identique. On remarque par exemple ici que ceci n'est pas tout à fait vrai pour les espèces 3 et 4, la branche menant à l'espèce 4 étant légèrement plus longue que celle menant à l'espèce 3 à partir du dernier nœud. Il s'agit de variations locales et minimes de l'horloge moléculaire qui doivent être négligées pour poursuivre. En contraignant la phylogénie à respecter strictement la constance de la vitesse évolutive, on obtient un arbre légèrement modifié (fig. 5.22).
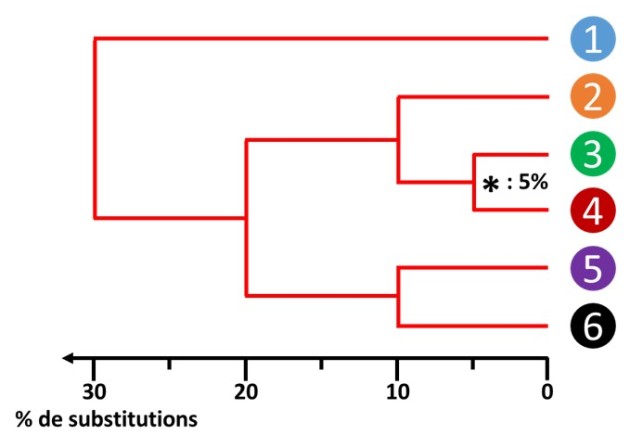
Il devient alors possible de positionner une échelle reflétant la différence constatée entre les séquences. On retrouvera par exemple Les 5% de différences mesurées entre les séquences des espèces 3 et 4.
La disponibilité de données paléobiologiques vont alors permettre de calibrer cet arbre. Il est par exemple connu que la séparation entre 3 et 4 date de 25 millions d'années. Suivant le principe de la constance de la vitesse évolutive, l'échelle des différences peut être substituée par une échelle de temps (fig. 5.23).
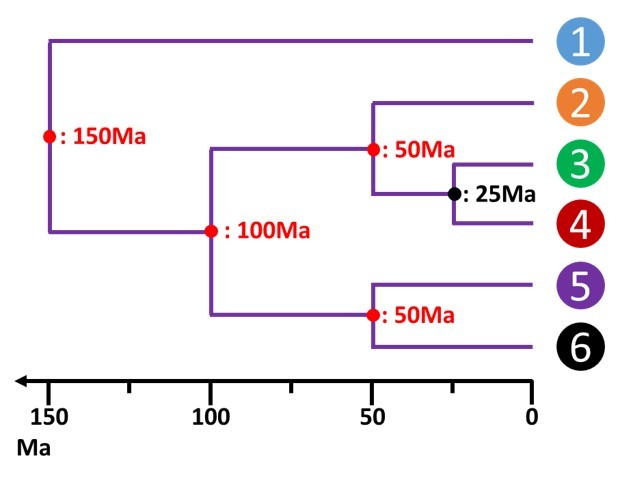
Chaque nœud, chaque séparation entre groupes taxonomiques peut alors être daté de façon absolue.
La datation est sujette à erreur
La datation paléontologique n'est cependant pas toujours précise. Une erreur d'approximation de la date de référence se répercute alors sur l'ensemble des dates déduites dans l'arbre. Dans l'exemple précédent, une évaluation erronée à 20Ma du dernier ancêtre entre les espèces 3 et 4 modifie l'échelle de la phylogénie (fig. 5.24).
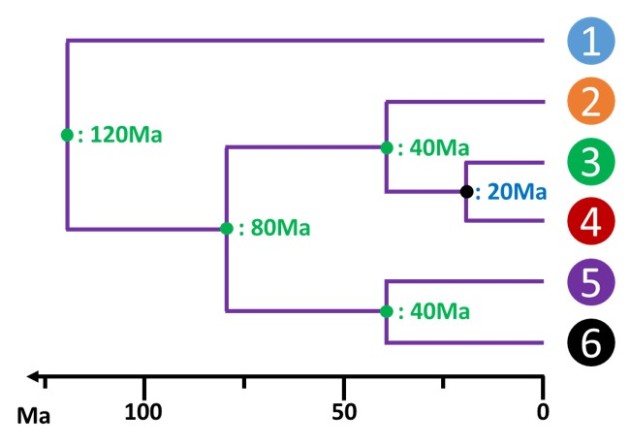
La datation est imprécise
Quelque soit la méthode de datation utilisée, le résultat s'accompagne d'une imprécision plus ou moins grande, sous forme d'un intervalle. Il est donc ensuite illusoire d'estimer précisément les autres dates associées à une phylogénie.
La parenté entre un fossile et des espèces modernes est alors parfois difficile à établir (fig. 5.25).
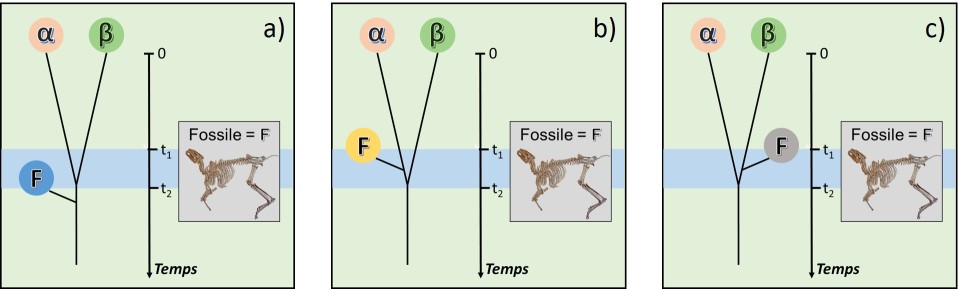
Si la datation du fossile propose une période entre t1 et t2, plusieurs hypothèses deviennent possibles :
F est identiquement apparenté à α et β (fig. 5.25a)
F est plus proche de α (fig. 5.25b)
F est plus proche de β (fig. 5.25c)