2.1.3 - La définition écologique
Définition : Ernst Mayr en 1982.
Une espèce est une communauté reproductive de populations, reproductivement isolée d'autres communautés et qui occupe une niche particulière dans la nature.
La niche écologique englobe l'ensemble des besoins requis par une espèce pour survivre dans son biotope et des actions réalisées pour les satisfaire (Blondel 1995 [1] pour de plus amples développements sur la notion de niche). La niche écologique doit être considérée comme un espace à n dimensions qui comporte la nature de la nourriture recherchée, le site et la période (du jour mais aussi de l'année) où cette nourriture est recherchée, les exigences de l'espèce pour sa reproduction... L'amplitude de la niche d'une espèce dépend des interations de cette espèce avec les autres espèces du peuplement. On peut donc distinguer une niche fondamentale, qui est la niche potentielle d'une espèce, et une niche réalisée, plus réduite, qui est la niche qu'elle occupe réellement dans la nature et qui résulte de la compétition avec les autres espèces.
Divers arguments justifient cette définition :
Les espèces biologiquement proches qui cohabitent dans un même milieu occupent des niches disjointes (fig. 2.5) ;
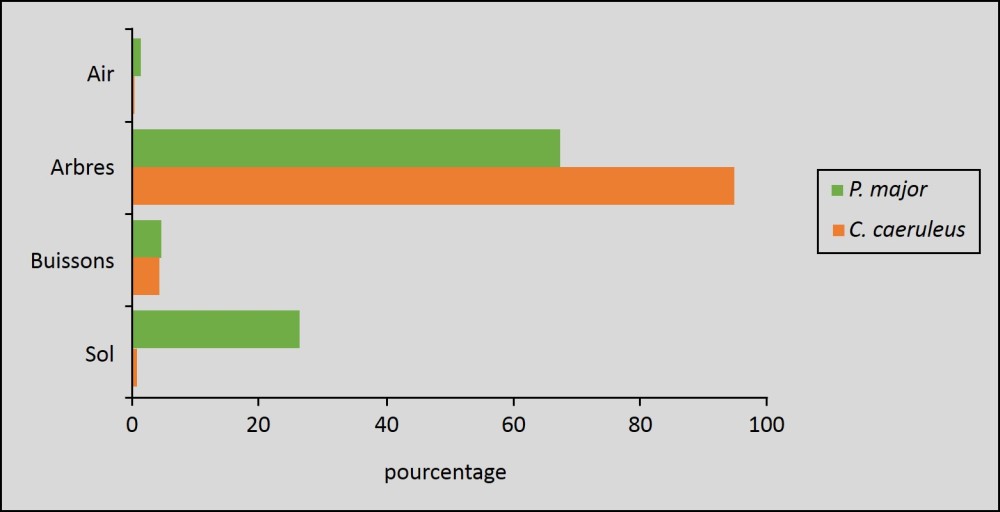
Sites de recherche alimentaire des mésanges bleue (Cyanistes caeruleus) et charbonnière (Parus major) en chênaie de chênes verts exprimé en % de temps passé sur chaque site (D'après Clamens 1988). Au sein des arbres la mésange bleue exploite les ramifications les plus fines des branches alors que la mésange charbonnière exploite plutôt le tronc.
Les déplacements de caractères d'espèces voisines en situation de sympatrie, exemple classique des pinsons de Darwin ;
Les introductions d'espèces qui entraînent la disparition d'autres espèces, cas de la disparition de l'écureuil roux en Grande-Bretagne suite à l'introduction de l'écureuil nord-américain ;
Les expériences de défaunation (expériences de Simberloff) (fig. 2.6) ;
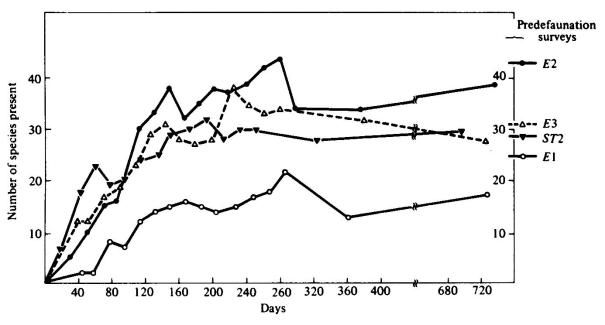
Reconstitution des peuplements en arthropodes d'un îlot au large de la Floride après défaunation expérimentale. Le nombre d'espèces s'accroît par colonisation, passe par un maximum puis se stabilise à un niveau proche de celui existant avant l'expérience . E1, E2, E3 et ST2 désignent les quatre îles défaunés.
Explication :
Simberloff et Wilson (1970 cités par Blondel 1995) ont détruit la faune d'arthropodes de certains îlots de Floride avec du methylbromide. Ils ont ensuite suivi la recolonisation de ces îlots par les arthropodes. Ils ont alors constaté que les îlots retrouvaient avec le temps un peuplement d'arthropodes semblable à celui d'avant l'expérience. Ainsi, les plus éloignés du continent, qui étaient les plus pauvres avant l'expérience, retrouvaient un nombre d'espèces plus faible que les plus proches du continent, qui étaient les plus riches en espèces, et qui retrouvaient pour leur part un nombre d'espèces élevé. Si les espèces rencontrées après recolonisation pouvaient être différentes de celles rencontrées avant l'expérience, les structures trophiques finales étaient identiques à celles observées avant destruction de la faune. Cependant, tous les îlots passaient par un stade caractérisé par un nombre d'espèces supérieur à celui observé avant l'expérience. Ceci suggère que le nombre d'espèces est un caractère du milieu étudié mais que par contre la nature des espèces qui l'occupe résulte du hasard et des aléas de la colonisation. Le stade plus riche en espèce que le stade initial, observé temporairement, correspond a un stade de déséquilibre. Si certaines espèces s'éteignent sans doute car le nombre de migrants est insuffisant, d'autres, arrivant ensemble et ayant les mêmes exigences écologiques, cohabitent tant que l'une n'élimine pas l'autre par compétition. Une fois atteint le niveau d'équilibre où toutes les niches sont occupées, le milieu est fermé et plus aucune espèce ne peut s'installer à moins qu'une autre ne s'éteigne.
Cette expérience est intéressante à plus d'un titre. Tout d'abord, elle montre que l'espace des ressources pris au sens large peut être divisé en un certain nombre d'unités discrètes qui n'accueillent chacune qu'une espèce. On peut donc caractériser une espèce par son mode d'utilisation du milieu, c'est à dire par sa niche écologique. ( D'après Allano L. & Clamens A.,[2])
Les crises géologiques qui libèrent des niches vides, comme la radiation des mammifères au Mésozoïque qui semble correspondre à l'occupation de niches laissées vides par les espèces disparues à la limite K-T ;
L'existence d'un isolement reproducteur entre espèces du même genre uniquement en condition de sympatrie.
Exemple : Répartition des espèces du genre Gilia
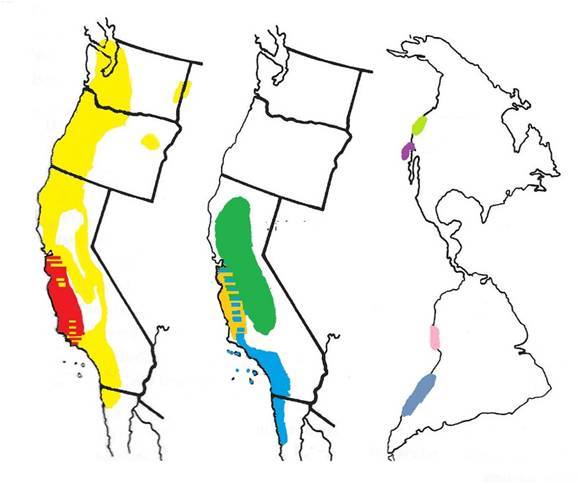
En 1966, Verne Grant[3] a étudié 9 espèces d'angiosperme du genre Gilia (famille des Polemoniaceae) (fig. 2.7), 5 étaient sympatriques avec des aires de répartition se chevauchant largement en Californie, les 4 autres étaient allopatriques dans des habitats maritimes d'Amérique du nord et du sud (fig. 2.8).
figure 2.7 : Gilia achilleifolia (Photo : National Parks Service (USA)) |
figure 2.8 : Carte de répartition des 5 espèces sympatriques (cartes de gauche) et des 4 espèces allopatriques (carte de droite) étudiées par Grant. Modifié d'après Briggs & Walters (2016)[4]. Rouge : G. clivorum, jaune : G. capitata, vert : G. tricolor, orange : G. achileaefolia, bleu : G. angelensis, bleu nuit : G. valdiviensis, rose : G. lomensis, violet : G. nevinii, vert clair : G. millefoliata. |

Il a alors réalisé des pollinisations croisées entre les espèces. Le résultat est donné dans le tableau ci-dessous.
Relations géographiques entre espèces | Nombre de combinaisons entre espèces pour la pollinisation | Nombre moyen de graines par fuit pour les différents croisements | Moyenne des moyennes |
|---|---|---|---|
Espèces sympatriques | 9 | 0,0 / 0,0 / 0,0 / 0,0 / 0,0 0,1 / 0,1 / 0,4 / 1,2 | 0,2 |
Espèces allopatriques | 5 | 7,7 / 16,7 / 19,6 / 21,9 / 24,9 | 18,1 |
Explication :
Les résultats de Grant[3] montrent que l'isolement reproducteurs n'existe qu'entre les espèces sympatriques, c'est-à-dire celles pour lesquelles la pollinisation croisée est possible et qui sont donc susceptible de s'hybrider. Cette particularité est connue sous le nom d'effet Wallace. L'isolement reproducteur constitue donc un trait d'histoire naturelle comme les autres soumis à la sélection naturelle.
Des études sur les drosophiles menées par Coyne et Orr (1989)[5] ont d'ailleurs montré qu'en situation d'allopatrie l'isolement postzygotique et l'isolement prézygotique évoluent à la même vitesse, alors qu'en situation de sympatrie l'isolement prézygotique, le moins couteux d'un point de vue énergétique, évolue plus vite. Il existe donc une pression de sélection conduisant à une diminution des flux de gènes entre espèces lorsque des espèces sont en situation de pouvoir échanger des gènes. Cette sélection a pour conséquence de maintenir les assortiments de gènes propres à chaque espèce et leur permettant d'exploiter une niche écologique particulière.