8.2.1- Rôle de Chordin et Noggin
Parmi les candidats moléculaires ayant un rôle dans la dorsalisation, Deux groupes d'interactions moléculaires interviennent. Ce sont BMP4/Noggin, Chordin, Follistatin et XWnt8/Frisbee, Goosecoid.
Ils sont tous produits et secrétés par les cellules de l'organisateur.


Ils apparaissent après la transition blastuléenne au stade 11 de la blastula et se renforcent au début de la gastrulation. Leur domaine d'expression se confond avec la lèvre dorsale du blastopore. Sur ces images, leurs transcrits sont mis en évidence par hybridation in situ, juste au niveau de la lèvre dorsale du blastopore, c'est à dire au niveau de l'organisateur de Spemann (Fig.213).
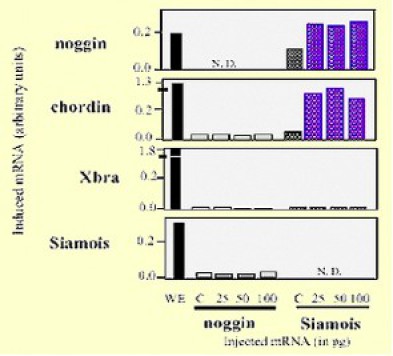
Leur expression est déclanchée par le facteur de transcription Siamois dont l'aire de répartition se confond ave le centre de Nieuwkoop et l'organisateur de Spemann. Des expériences de surexpression ectopique Siamois montrent que l'expression de siamois est suivi dans le clone où il s'exprime, de l'expression de Noggin et Chordin (Fig.214).Les expériences contrôles négatives avec la surexpression de Noggin n'entraîne l'expression ni de Chordin ni de Noggin. XBra et Siamois ne sont pas non plus régulés par Siamois.
Noggin et Chordin sont régulés positivement par les facteurs de croissance Xnr 1, 2 et 4 (mais pas Xnr3). La polarisation dorsale, avec la caténine et l'induction mésodermique avec VegT régulent donc, en synergie, l'expression de Noggin et Chordin.
Noggin et Chordin sont des antagonistes du facteur de croissance TGF : BMP4 (Bone Morphogenetic Protein). La liaison moléculaire est directe.
BMP4 est une molécule répartie dans tout l'embryon, et possède un effet ventralisant. C'est donc dans la région dorsale que Chordin et Noggin vont inhiber BMP4. L'effet inhibiteur est majoritaire car, par exemple, la concentration de Chordin est de 1000 à 10000 fois supérieure à celle de BMP4 soit de 6 à 12mM pour 1pmole.
Cependant, chaque molécule doit avoir un rôle différent puisque les constantes d'affinité sont différentes, soit 300pM pour Noggin et 20pM pour Chordin. Noggin possède donc 15 fois plus d'affinité pour BMP4 que Chordin.
Des expériences montrent que BMP4 possède un rôle basique de ventralisation qui est inhibé dans la région dorsale par Noggin et Chordin.
Les antisens de Chordin ou Noggin entraînent l'expansion du mésoderme de type ventral dans toute la zone marginale y compris dorsale.
Enfin, la surexpression des ARNm de BMP4, ou bien l'injection de la protéine BMP4 ventralisent l'embryon. On obtient alors un phénotype semblable à celui obtenu par l'irradiation du pôle végétatif après la fécondation.
La dorsalisation s'explique donc par une compétition entre deux molécules morphogènes, l'une ubiquiste, BMP4, l'autre restreinte à la DMZ, secrétée à partir de l'organisateur de Spemann, qui inhibe la fonction de la première en interagissant avec elle (Fig.215).
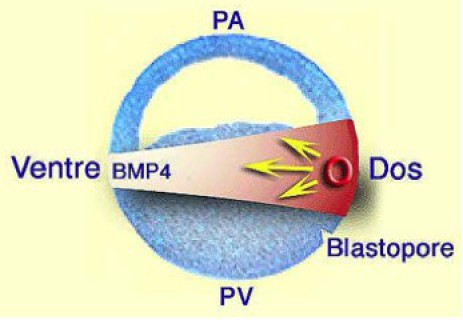
Cette interaction s'effectue de la manière suivante (Fig.216):
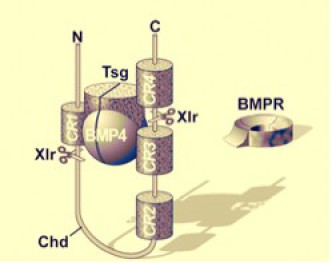
BMP4 est associé à une protéine, produit du gène Twistgastrulation (Tsg). C'est ainsi que BMP4 est naturellement présenté à son récepteur BMPR
Chordin se lie à ce complexe au niveau des unités répétitives CR1 à CR4.
Ainsi le complexe Chordin – BMP4 – Tsg empêche BMP4 de se lier à son récepteur transmembranaire d'où l'inhibition de BMP4.
Mais il existe un mécanisme de régulation négative de Chordin dans la mesure où une autre protéine appelée Xolloïd reconnaît le complexe Chordin – BMP4 – Tsg, coupe la boucle peptidique de Chordin comprise entre les unités répétitives CR1 et CR4 et libère ainsi BMP4 – Tsg qui redevient actif.
Ceci explique pourquoi la concentration en Chordin et Noggin doit être bien supérieure à celle de BMP4 pour que la dorsalisation ait lieu.
Au total, l'organisateur secrète les antagonistes de BMP4 tout au long de la gastrulation.
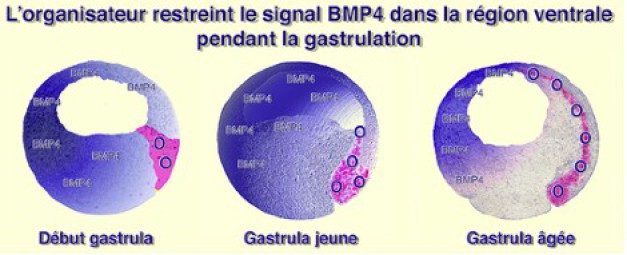
Or, le territoire de la lèvre dorsale du blastopore s'invagine, migre sur le plafond du blastocoele, se déforme et s'allonge dans le sens antéropostérieur. Les antagonistes de BMP4 inhibent les territoires qu'ils rencontrent à cette occasion. Ce sont les tissus mésodermiques latérodorsaux et l'ectoderme dorsal.
Le schéma ci-dessus rend compte du déplacement et de la déformation de l'organisateur au cours de son invagination (Fig.217).
Vers la fin de la gastrulation le domaine d'expression de BMP4 se restreint aux régions latéroventrales, à l'exclusion de la face dorsale où se différencient le système nerveux, la chorde et les somites (Fig.218).
De nombreuses expériences permettent de démontrer l'alternative BMP4/Chordin Noggin.
Par exemple (Fig.219):
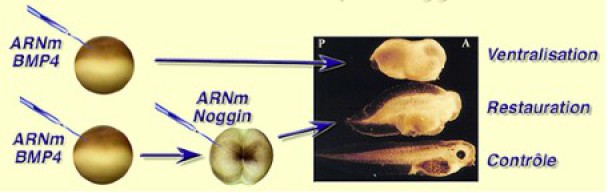
L'injection des ARNm de BMP4 dans un œuf fécondé ventralise l'embryon.
La co-injection ou la réinjection des ARNm de Noggin au stade 2 cellules corrige l'effet ventralisant de BMP4 et restaure une partie des structures dorsales.