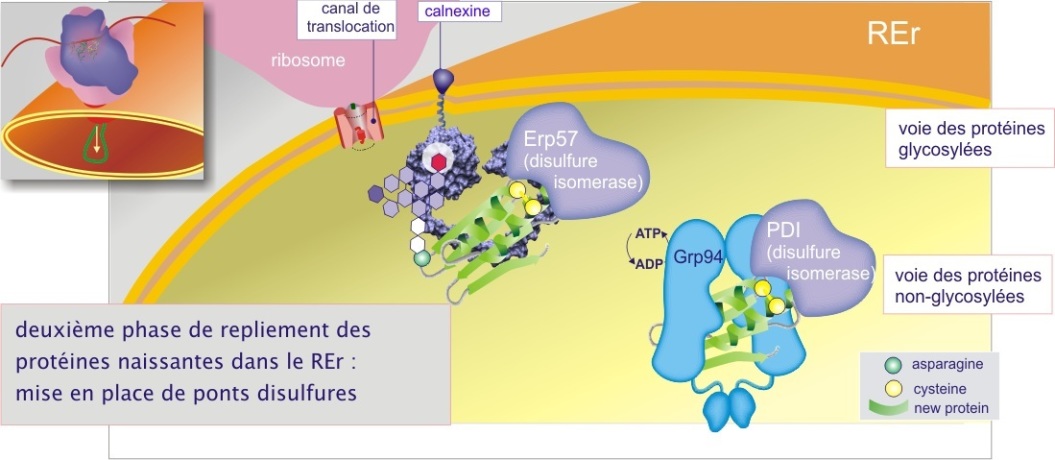
Les ponts disulfures
Contrairement au cytosol le RE constitue un environnement oxydant capable d'arracher des électrons (et aussi par conséquent, des protons) à la protéine nouvellement synthétisée. Un tel environnement favorise la formation de ponts disulfures entre les cystéines dans la réaction suivante :
\(\rm dithiol\ SH\ SH+\frac 12O_2\to disulfures\ S-S+H_2O\ (oxydation)\)
Les ponts disulfures ont la propriété de stabiliser la protéine en réduisant son entropie configurationnelle dans l'état non replié. Plus tard, selon l'environnement, leur mise en place ou leur coupure va établir un contrôle, qualifié de contrôle rédox, de l'activité de la protéine (voir figure 7B).
La réaction d'oxydation est rendue possible par la présence de FAD (flavine adénine dinucléotide), accepteur d'électrons (et de protons) du RE. Le principe général veut que les électrons d'une structure moléculaire possédant un bas potentiel rédox vont vers une structure possédant un haut potentiel rédox (voir aussi : ). Les différentes protéines impliquées dans la formation de ponts disulfures possèdent toutes le pli de « thirédoxine » (domaine Trx) contenant un site catalytique composé de deux cystéines dans un motif –C–X–X–C–. Le potentiel rédox du site catalytique varie selon les protéines qui le portent, ce qui détermine leur place dans la chaîne des événements et leur choix de substrats.
Les évènements de cette chaîne sont les suivants : la protéine cliente interagit avec une disulfure isomérase, soit ERp57 (endoplasmic reticulum protein de 57 kDa) dans le cas de protéines liées aux lectines chaperonnes, soit PDI (protein disulfure isomerase de 59 kDa (aussi connu comme ERp59)) dans le cas de protéines liées à d'autres chaperonnes (telles que BiP ou Grp94). La disulfure isomérase oxyde les groupements SH (dithiol) en arrachant deux électrons et deux protons et crée donc un pont disulfure (voir figure 8). Les électrons et protons arrachés sont transitoirement hébergés par les disulfides (S–S) du site catalytique formant ainsi les dithiols (SH SH). Une deuxième protéine, qualifiée d'oxyréductine, porteuse elle aussi du site catalytique (–C–X–X–C–), oxyde le PDI ou l'ERp57 et transporte les électrons et protons vers le FAD, le convertissant en FADH2. L'oxyréductine de PDI est Ero1–L, celle de l'Erp57 n'a pas encore été identifiée. La séquence peut recommencer lorsque le FADH2 aura cédé ses électrons et protons à l'oxygène, pour donner de l'eau (voir figure 8).
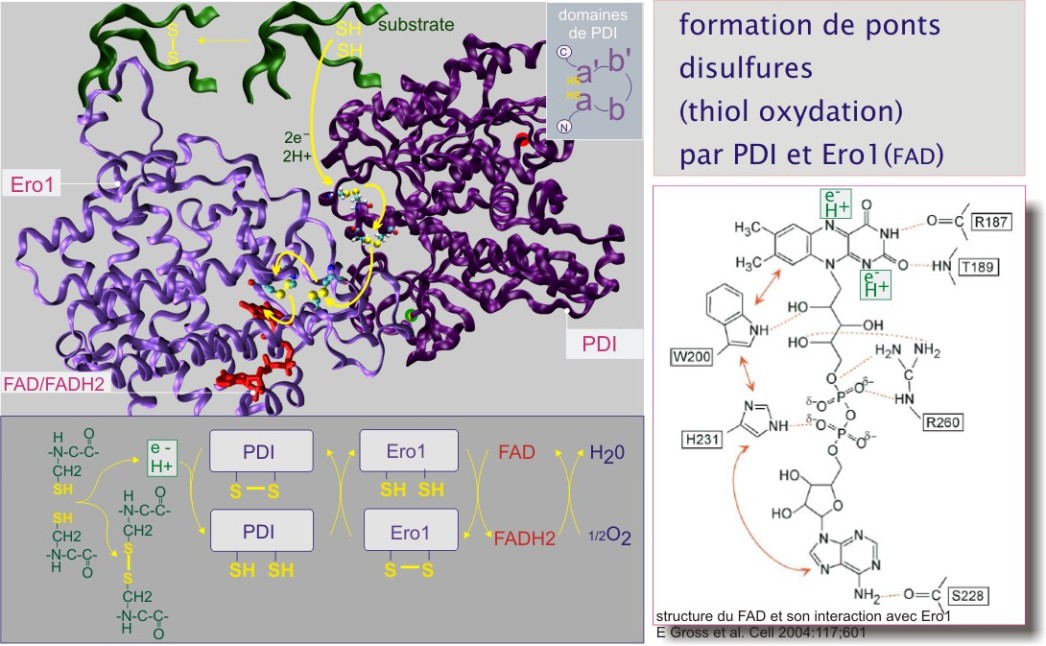
Après que les premiers ponts disulfures aient été formés, les disulfures isomérases continuent d'aider la protéine à se plier en favorisant l'échange interne de ponts disulfures (les premiers ponts établis n'étant pas forcément les ponts définitifs de la protéine native). Pendant ces réactions d'isomérisation, la disulfure isomérase est transitoirement liée au substrat par une liaison covalente (S–S).