Le rôle des molécules d'adhérence dans la cohésion tissulaire
Introduction
La structure tissulaire est activement maintenue par un principe d'affinité entre cellules. Les molécules d'adhérence forment la base de cette affinité spécifique. Les tissus conjonctifs d'une part, et épithéliaux d'autre part, représentent deux extrêmes dans lesquels la matrice et les systèmes d'adhérence intercellulaire jouent des rôles structuraux complètement différents.
Dans les tissus conjonctifs, la matrice extracellulaire est abondante et supporte la plus grande partie des tensions auxquelles les tissus sont soumis. Les interactions entre cellules sont limitées, du fait de leur distribution lâche. Dans les épithéliums qui tapissent toutes les cavités et les surfaces externes du corps (et également dans les endothéliums), les cellules sont étroitement associées en feuillets. La matrice extracellulaire est peu abondante, constituant une couche fine, la lame basale, sous-jacente aux cellules. Les cellules elles-mêmes, plutôt que la matrice, subissent la plus grande partie des tensions. C'est grâce aux jonctions cellulaires spécialisées que l'épithélium et l'endothélium sont suffisamment résistants pour constituer une barrière capable de séparer deux compartiments ().
Définition : Epithélium et endothélium :
L'épithélium peut se définir comme un assemblage de cellules (en couches plus au moins nombreuses) qui constitue une barrière entre l'organisme et son environnement. Exemples : la peau, la muqueuse de la vessie et du tube digestif sont recouvertes par des épithéliums.
L'endothélium est le revêtement interne monocouche des vaisseaux sanguins et lymphatiques, et constitue une barrière entre deux compartiments corporels.
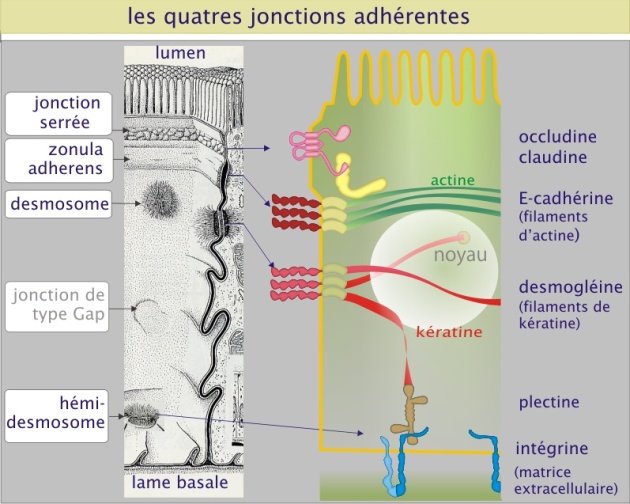
Les jonctions intercellulaires des cellules épithéliales peuvent être classées en trois groupes (selon leur ultrastructure et leur fonction) :
les jonctions étanches, « jonctions serrées » ou « zonula occludens » capables de limiter la perméabilité de l'épithélium (ou de l'endothélium),
les jonctions d'ancrage, « zonula adherens » et « desmosomes », qui permettent l'attachement mécanique des cellules entre elles,
les jonctions communicantes, « jonctions de type gap », qui permettent le passage de signaux chimiques ou électriques entre les cellules.
Les jonctions serrées
L'étanchéité de l'épithélium et de l'endothélium est assurée par les jonctions serrées. Elles constituent un rapprochement étroit et localisé des membranes de deux cellules voisines qui limite considérablement le passage des solutés par l'espace intercellulaire (barrière paracellulaire). Cette barrière oblige le soluté à transiter au travers du feuillet cellulaire grâce à des transporteurs membranaires sélectifs (transport transcellulaire). La jonction n'est pas entièrement étanche : selon la nature du tissu, certains solutés peuvent ou non la franchir. La jonction est due à des interactions homophiles qui concernent plusieurs molécules d'adhérence. La première est l'occludine. (latin occludere = enfermer), protéine transmembranaire d'un poids de 64 kDa, constituée d'une chaîne polypeptidique qui traverse quatre fois la membrane. La deuxième est la claudine (latin claudere = fermer), protéine transmembranaire d'un poids de 22 kDa, constituée également d'une chaîne polypeptidique qui traverse quatre fois la membrane. La claudine fait partie d'une famille de 20 membres, dont plusieurs peuvent être présents dans une même jonction. Les nombreuses possibilités de combinaison des claudines pourraient expliquer les différences dans la perméabilité des jonctions serrées selon les tissus. La troisième protéine est JAM (junctional adhesion molecule), une protéine d'un poids moléculaire de 33 kDa et dont la chaîne polypeptidique traverse la membrane une seule fois.
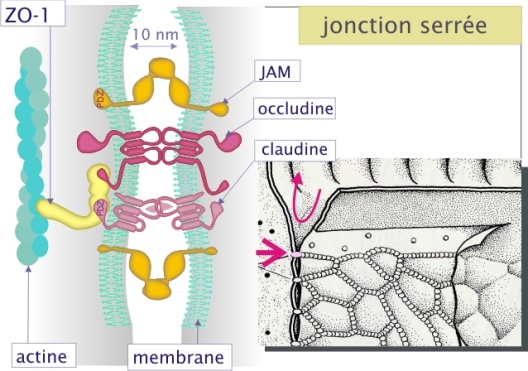
Occludine, claudines et JAM s'assemblent pour former un réseau de fibrilles qui ceinture le domaine apical des cellules épithéliales (ou le domaine membranaire latéral des cellules endothéliales). Les jonctions serrées sont liées avec des protéines intracellulaires comme ZO-1 et ZO-2 (zonula occludens) qui à leur tour sont liées au cytosquelette (actine). Cette interaction (avec le cytosquelette) détermine la localisation de la jonction au domaine apical de la cellule.
Remarque :
Les claudines 3 et 4 peuvent se comporter comme des récepteurs d'une entérotoxine bactérienne (CPE) produite par Clostridium perfringens. La nourriture contaminée par cette bactérie cause une diarrhée due à une augmentation de la perméabilité paracellulaire et une perturbation profonde de la réabsorption aqueuse au niveau du tube digestif. Au départ, c'est la liaison de la toxine qui fait disparaître les claudines et change profondément la structure des fibrilles des jonctions serrées des entérocytes.
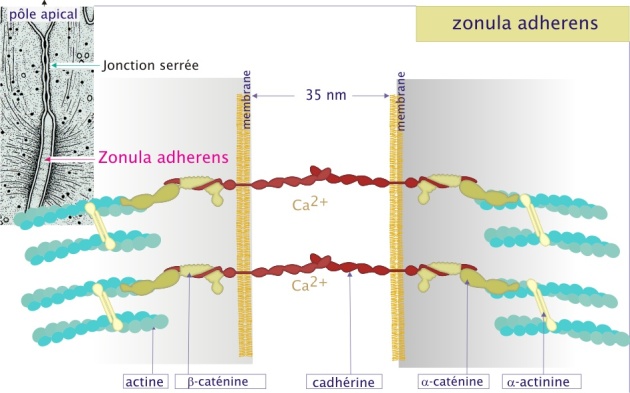
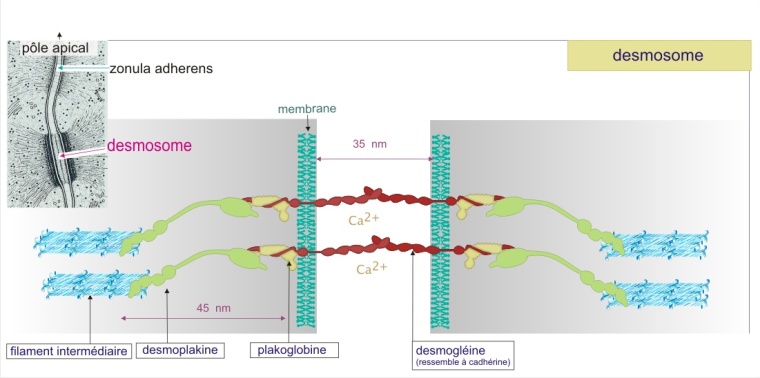
Les zonula adherens et les desmosomes
Ces jonctions d'ancrage permettent à des groupes de cellules d'agir comme des unités structurales solides en associant les éléments du cytosquelette d'une cellule à ceux d'une autre. Les zonula adherens sont des sites de liaison pour les filaments d'actine. Les desmosomes (du grec de desmos : lien) sont des sites de liaison pour les filaments intermédiaires (kératine dans les cellules épithéliales). Les deux types de jonctions sont composés de molécules transmembranaires d'adhérence : les cadhérines. Nous avons déjà vu les cadhérines quand nous avons parlé des interactions entre cellules dans l'assemblage des tissus ; ici nous voyons que les cadhérines sont aussi des constituants de structures complexes et définitives (que l'on peut voir en microscopie électronique).
Dans le cas des zonula adherens la cadhérine (forme E-cadhérine) est d'abord liée à la -caténine, protéine de 80 kDa, qui sert donc d'intermédiaire entre elle et l'actine. Les zonula adherens connectées au cytosquelette forment la ceinture d'adhérence dans le feuillet épithélial. Cette ceinture renforce l'intégrité tissulaire (voir l'animation de la figure 12 ci-dessous).
![]()
Macromedia Flash - 40Ko
Les desmosomes contiennent aussi de la cadhérine sous forme de desmogléine (poids moléculaire ~160 kDa) et de desmocolline (poids moléculaire ~110 kDa), liée cette fois à la plakoglobine (une protéine de 110 kDa qui ressemble à \(\beta\)-caténine), elle-même liée à la desmoplakine (d'un poids moléculaire de 285 kDa ) qui fixe les filaments intermédiaires. Les filaments intermédiaires forment une charpente structurale pour le cytoplasme et assurent sa résistance aux forces latérales (voir l'animation de la figure 13 ci-dessous).
![]()
Macromedia Flash - 36Ko
Exemple :
Un exemple montre l'importance de ces complexes jonctionnels étroitement liés au cytosquelette. Dans les tumeurs épithéliales les cadhérines perdent leur fonction et, les cellules ne construisant plus de jonctions intercellulaires solides, sont plus sensibles aux signaux de prolifération (formation de polypes) et deviennent susceptibles de migrer et donc d'envahir d'autres tissus (processus métastatique).
Les jonctions de type gap
Cliquez pour revoir la communication entre cellules, abordée dans la ressource « Transport membranaire »