Le rôle des molécules d'adhérence dans la circulation des cellules immunitaires
Les sélectines, intégrines, CAM et la migration transendothéliale
Les molécules d'adhérence sont également à la base des processus qui permettent la circulation des cellules du système immunitaire dans l'ensemble des organes. Le phénomène d'inflammation localisé au site de lésion ou d'infection attire les cellules immunitaires compétentes (les leucocytes tels que monocytes, lymphocytes et granulocytes) nécessaires au déclenchement de la réparation tissulaire et de la lutte contre les agents pathogènes. En effet, au niveau du site d'inflammation, les leucocytes quittent la circulation sanguine et s'infiltrent dans le tissu enflammé.
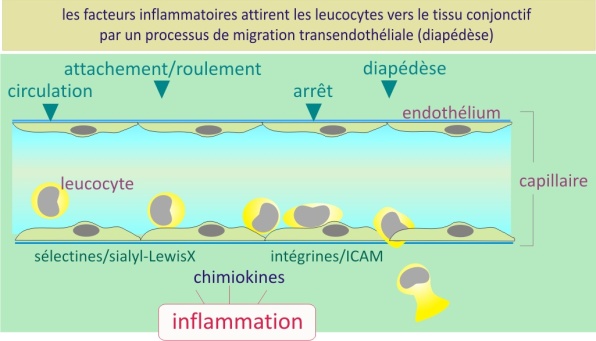
Ce processus débute par une liaison faible entre les leucocytes et les cellules endothéliales (vasculaires), liaison où interviennent les sélectines (E- et P-sélectine) et les motifs glucidiques (jouant le rôle de ligands comme sialyl LewisX). Dans les circonstances normales, cette faible interaction, combinée au flux sanguin, résulte en un mouvement de roulement des leucocytes à la surface de l'endothélium.

![]()
Macromedia Flash - 30Ko
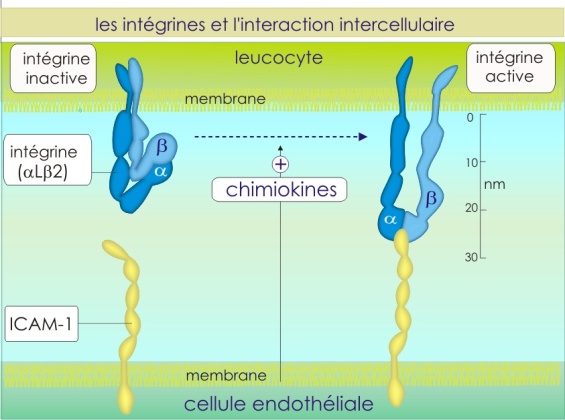
Cependant, au site de l'inflammation, les cellules endothéliales relarguent des chémokines. Ces molécules ont pour rôle d'immobiliser les leucocytes roulants et de favoriser leur traversée de l'endothélium. Pour cela elles activent, grâce à un signal intracellulaire, les intégrines, autres molécules d'adhérence présentes à la surface des leucocytes. Les intégrines (\(\alpha L \beta2\)) activées se fixent sur les I-CAM-1 (qui sont fortement exprimées sur l'endothélium au site de l'inflammation) provoquant ainsi une immobilisation totale du leucocyte. En se déformant, celui-ci migre vers le tissu en passant entre les cellules endothéliales. Une fois dans le tissu, les leucocytes assurent deux rôles ; reconstruire le tissu endommagé et en cas de lésion septique, éliminer les agents pathogènes. L'accumulation des leucocytes est la résultante de deux phénomènes : surexpression des sélectines et I-CAMs sur les cellules endothéliales et relargage des chémokines par ces cellules uniquement au site de l'inflammation.
![]()
Macromedia Flash - 60Ko
Il existe plusieurs sélectines (par exemple sélectine-E, -P et -L d'un poids moléculaire d'environ 90 kDa) qui interagissent avec les motifs glucidiques présents sur plusieurs ligands. Il existe également plusieurs I-CAMs (formes 1-3 d'un poids moléculaire compris entre 29 et 55 kDa). Les I-CAMs établissent des interactions hétérophiles avec les intégrines.
Les intégrines forment une grande famille de molécules d'adhérence. On les trouve sous forme d'hétérodimères composés de sous-unités \(\alpha\) et \(\beta\) (poids moléculaire d'environ 120 kDa et 90 kDa) qui elles-mêmes présentent plusieurs types susceptibles de former diverses combinaisons (\(\alpha 5 \beta 1\) ou \(\alpha L \beta 2\) etc.) Sur les leucocytes mais aussi sur les plaquettes circulant dans le sang, les intégrines se trouvent dans un état inactif (non reconnaissance de leur ligand). Leur activation nécessitera la présence de chémokines. En revanche, dans le cas des cellules assemblées en tissu, les intégrines sont toujours dans un état actif et permettent donc le bon ancrage cellulaire (voir la section sur les molécules d'adhérence interagissant avec la matrice extracellulaire, ainsi que la ).