Le rôle des molécules d'adhérence dans l'assemblage cellulaire
Les molécules d'adhérence sont à la base du mécanisme qui permet à une cellule embryonnaire de s'attacher au bon moment à une cellule qu'elle aura identifiée (formant ainsi un tissu puis un organe).
Les molécules d'adhérence intercellulaire CAM dans développement de la rétine embryonnaire
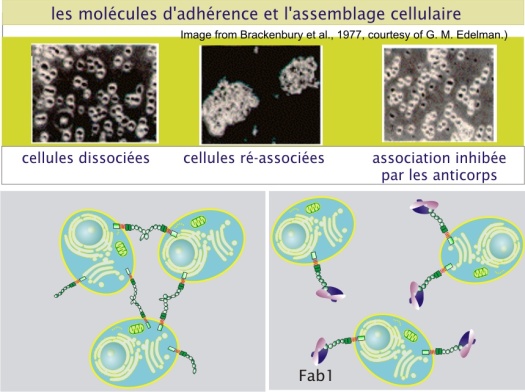
Une expérience déjà ancienne démontre le rôle des molécules d'adhérence dans l'assemblage des tissus : des cellules de la rétine embryonnaire de poulet (stade embryonnaire de 10 jours) expérimentalement dissociées se réassocient spontanément en environ 30 minutes en amas cellulaires lorsqu'elles sont mises en culture. Certains anticorps (plus précisément leurs fragments Fab1), dirigés contre des molécules membranaires de ces cellules sont capables d'inhiber ce réassemblage.
Cliquez pour plus de détail sur l'isolement des fragments d'IgG
Après une procédure de purification, ces anticorps se sont avérés reconnaître une protéine transmembranaire d'un poids moléculaire de 140 kDa, qu'on appelle « neural cellular adhesion molecule » (N-CAM) qui joue donc un rôle dans la reconnaissance des cellules entre elles. Les N-CAMs établissent des interactions homophiles : les N-CAMs d'une cellule se lient aux N-CAMs des cellules adjacentes (voir figure 4 ci-dessous).
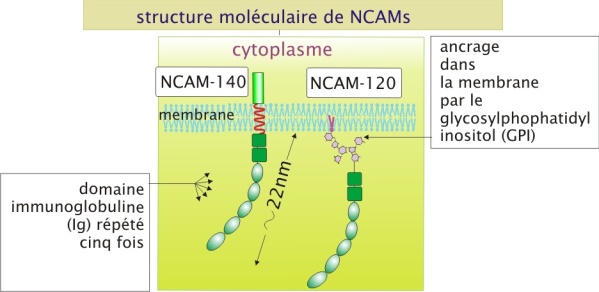
La protéine N-CAM est codée par un seul gène mais grâce aux épissages alternatifs de l'ARNm elle peut se présenter sous trois formes :
180 et 140 kDa, qui sont des protéines transmembranaires dont la chaîne polypeptidique traverse la membrane une seule fois en hélice-\(\alpha\)
et une troisième forme de 120 kDa qui est ancrée dans la membrane par un lipide (protéine membranaire intrinsèque).
En raison de sa structure moléculaire, la N-CAM est membre d'une famille de molécules d'adhérence qui appartient à la superfamille des immunoglobulines (anticorps). Cette famille comprend d'autres membres tels que I-CAM-1,2,3 et V-CAM (voir le paragraphe « circulation des cellules immunitaires »). Bien que l'expression de N-CAM ait d'abord été démontrée dans les neurones, elle a été retrouvée ailleurs, par exemple dans les cellules musculaires et les lymphocytes.
Les molécules d'adhérence dans la compaction embryonaire
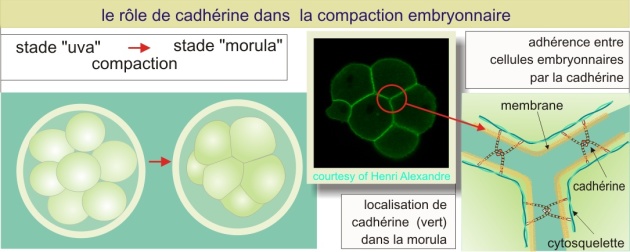
Dans le développement embryonnaire de la souris (par exemple), il y a un moment ou l'embryon se compacte : il change de structure en passant du stade « uva » au stade « morula » (du latin uva -> raisin et morula -> mûre).
Cliquez pour plus de détail sur le processus de compaction
![]()
Macromedia Flash - 15Ko
Ce processus est dépendant des molécules d'adhérence et de la présence de Ca2+. Comme précédemment, des anticorps reconnaissant certaines molécules présentes sur la membrane cellulaire sont capables d'inhiber cette transition. La molécule d'adhérence impliquée dans ce processus est la E-cadhérine (Ca2+- dependent adhesion). Les cadhérines jouent donc un rôle important dans la reconnaissance sélective des cellules embryonnaires et également dans l'établissement de leur polarité cellulaire. Les cadhérines sont des molécules d'adhérence dont la chaîne polypeptidique passe la membrane une seule fois. Elles ont un poids moléculaire d'environ 80 kDa. Comme les CAMs elles sont constituées de cinq modules répétitifs (les domaines Cad), avec la fixation de trois ions de Ca2+ au niveau de chaque interface de domaine. Les cadhérines se lient entre elles selon le mode homophile. Elles forment une famille comprenant 12 membres (à ce jour). A l'intérieur de la cellule, les cadhérines sont reliées au cytosquelette par deux intermédiaires : la \(\beta\)-caténine (poids moléculaire de 85 kDa) et l'alpha-caténine (poids moléculaire de 100 kDa) (voir aussi « la zonula adherens » dans cette ressource). La fixation au cytosquelette renforce la liaison cellule-cellule.
