La ribulose bisphosphate carboxylase-oxygénase
Apport scientifique : nature, structure de l'enzyme
L'enzyme clef du cycle de Calvin-Benson-Bassham est la RuBisCO (EC 4.1.1.39; http://www.ebi.ac.uk/intenz/query?cmd=SearchID&id=10762&view=SIB) car son abondance et son efficacité déterminent la vitesse avec laquelle le CO2 est fixé par les chloroplastes et donc le taux de croissance d'une plante. La RuBisCO, qui peut représenter jusqu'à 50% des protéines totales chez les plantes, serait l'enzyme la plus abondante de la planète Terre. Etant donné son rôle central dans la production primaire, la RuBisCO est une enzyme très largement étudiée et sa structure tridimentionnelle est maintenant bien connue (Figure 46). La structure de la RuBisCO diffère légèrement entre les plantes supérieures et les algues. La RuBisCO est une grosse enzyme de 70 000 Da. Elle est formée de quatre hétérodimères (Figure 46). Chaque hétérodimère est composé d'une grande sous-unité (représentée en nuances de gris dans la figure 46) et d'une petite sous-unité (représentée en jaune et rouge dans la figure 46). Comme vous le constatez, l'enzyme présente une symétrie d'ordre 4. L'insert dans la figure 46 représente la RuBisCO de l'épinard, pour laquelle on constate de légères différences dans la petite sous-unité. Chaque hétérodimère contient du zinc, du magnésium et possède un site actif. C'est ce site actif qui est utilisé pour transformer une molécule de RuBP en 2 molécules de PGA (Figure 45).
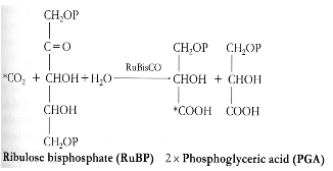
A. Organisation globale de la RuBisCO : L'enzyme est formée de quatre hétérodimères, chacun étant composé d'une grande sous-unité (représentée en nuances de gris) et d'une petite sous-unité (représentée en jaune et rouge). La structure globale présente une symétrie d'ordre 4. L'insert dans la figure représente la RuBisCO de l'épinard, pour laquelle on constate de légères différences dans la petite sous-unité.
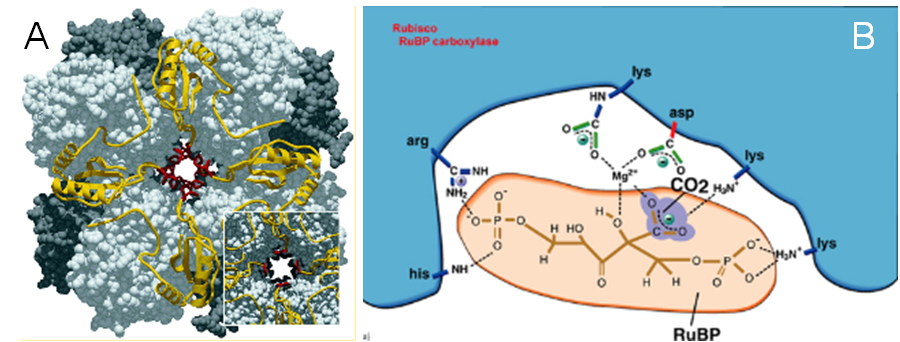
B. Schéma d'un site actif de la RuBisCO avec son substrat, le RuBP en place est en train d'être carboxylé.
Apport scientifique : fonction de l'enzyme - La photorespiration |
Jusqu'ici, nous avons parlé de la fonction “carboxylase” de la RuBisCO mais comme son nom vous l'avait laissé percevoir, cette enzyme peut exercer une autre activité enzymatique : l'oxygénation. Il est remarquable que cette activité se déroule dans le même site actif que la carboxylation. Ceci signifie que les deux activités sont en compétition et que l'activité qui sera dominante dépend de la concentration relative en substrats à proximité de l'enzyme. L'activité d'oxygénation se produit à la lumière quand le rapport O2/CO2 est élevé à proximité de la RuBisCO. Cette situation se produit chez les plantes terrestres quand celles-ci ferment leurs stomates (Figure 47).
Cellules de l'épiderme foliaire de polypode avec 2 stomates (X 150) . christohpe Carillon. . Disponible sur : http://acces.ens-lyon.fr/evolution/evolution/relations-de-parente/enseigner/activites-pratiques-et-classification/les-tp/tp-presence-de-stomates
Comme vous l'avez découvert au premier semestre, la fermeture des stomates se déroule lors d'une élévation de la température de l'air, ce qui permet aux plantes de ne pas trop perdre d'eau. Dans ces conditions, l'approvisionnement en CO2 est très limité alors que la phase claire de la photosynthèse continue à produire de l'O2 qui par ailleurs ne peut pas s'échapper puisque les stomates sont fermés. Le rapport des pressions partielles pO2/pCO2 s'accroît autour de la RuBisCO, ce qui enclenche la fonction oxygénase de l'enzyme (Figure 47). Donc, lorsque c'est le CO2 qui domine, l'activité de carboxylation se déroule mais lorsque c'est l'O2, c'est l'activité d'oxydation qui se réalise. Voyons maintenant les conséquences de cette réaction. Une molécule de RuBP fixe une molécule O2 au lieu d'une molécule de CO2 puis elle est scindée en une molécule de PGA (3 carbones) et une molécule de 2-phosphoglycolate (2 carbones) qui est immédiatement déphosphorylée pour donner une molécule de glycolate (Figure 47). Cette molécule est transportée dans le peroxysome. Dans ce compartiment le glycolate est progressivement transformé en glycine. Cette voie métabolique requiert la présence d'O2 et forme du peroxyde d'oxygène (H2O2). Il s'agit d'une molécule présentant un fort pouvoir oxydatif qui ne doit donc pas s'accumuler. Pour éviter cela, les peroxysomes contiennent un arsenal enzymatique, dont la catalase, très abondante dans le peroxysome, permettant l'élimination d'H2O2. C'est la présence de grandes quantités d'enzymes formant et dégradant le peroxyde d'hydrogène qui a conduit Christian DE DUVE à appeler ce compartiment cellulaire : peroxysome. La glycine est transportée dans la mitochondrie où elle est transformée en sérine. La réaction génère de l'ammoniac et une molécule de dioxyde de carbone. Cette réaction qui semble anodine a pourtant une importance considérable car lorsqu'elle se déroule, les plantes perdent du carbone. Nous reviendrons sur cet aspect un peu plus loin. La sérine est importée dans le peroxysome où elle est transformée en glycérate. Le glycérate est lui-même importé dans le chloroplaste où il est phosphorylé en PGA qui peut entrer à nouveau dans le cycle de Calvin-Benson-Bassham (Figure 47). Si vous faites abstraction du détail des réactions présentées dans la figure 47 et du contexte cellulaire dans lequel elles se déroulent, vous constaterez qu'elles nécessitent la prise d'oxygène et la libération de dioxyde de carbone. C'est pour cette raison que cet enchaînement de réactions porte le nom de photorespiration. Bien entendu, les plantes ne respirent pas de cette manière. Comme la majorité des organismes, les plantes respirent grâce à leurs mitochondries.
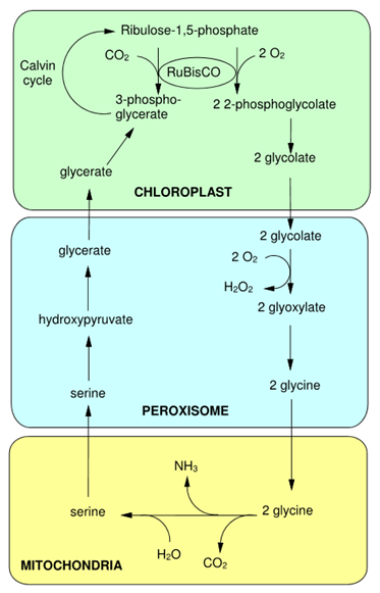
La photorespiration est la conséquence de la fonction “oxygénase” de la RuBisCO. Elle implique des réactions localisées dans le chloroplaste, la mitochondrie et le peroxysome. Du dioxygène est fixé par la RuBisCO tandis que du CO2 et de l'H2O2 sont produits pendant la transformation du glycolate en glyoxylate. Deux acides aminés importants, la glycine et la sérine, servent d'intermédiaires entre le peroxysome et la mitochondrie. La glycine est métabolisée en sérine dans ce dernier compartiment, ce qui génère du CO2. Le bilan de la photorespiration est donc une molécule de CO2 produite pour deux molécules d'O2 utilisées.
Rappelons-nous que le CO2, qui est fixé par la RuBisCO, provient de l'air arrivant par les stomates. Or dans l'atmosphère, il y a environ 500 fois plus d'O2 que de CO2. Cependant, la RuBisCO, ne fonctionne pas dans l'air mais dans le stroma des chloroplastes. Or celui-ci est aqueux et dans l'eau, le coefficient de dissolution de chacun des deux gaz est tel que l'O2 n'est plus qu'environ 25 fois plus abondant que le CO2. Malgré cela, il est peu probable que la réaction de carboxylation se déroule sauf si la sélectivité de l'enzyme pour ces deux gaz est très différente. C'est en fait ce qui est mesuré : selon les espèces, la sélectivité pour le CO2 est entre 20 et 280 fois plus importante que pour l'O2 (Griffiths 2006). Les plantes qui utilisent le cycle de Calvin pour fixer directement le CO2 sont appelées “plantes en C3” en raison de l'intermédiaire à trois carbones, le PGA, formé à chaque tour de cycle. Pour alimenter le chloroplaste en CO2, les plantes en C3 doivent maintenir leurs stomates ouverts, ce qui implique une évapotranspiration importante. Elles perdent jusqu'à 97 % de l'eau absorbée par les racines (Raven & Edwards 2001). Sous nos latitudes, ces plantes peuvent proliférer car les eaux souterraines sont abondantes et, l'intensité de l'éclairement solaire et la température restent modérés. Si une carence en eau se fait jour, les plantes en C3 ferment leurs stomates pour limiter la perte d'eau, ce qui a pour conséquence de réduire l'apport en CO2 alors que la quantité d'O2 augmente en raison de la photosynthèse. Dans ces conditions, l'activité “oxygénase” de la RuBisCO prend le pas sur l'activité “carboxylase” et la photorespiration se met en place.
Comment les plantes ont-elles colonisé les milieux secs ? C'est que nous allons découvrir bientôt.