2.2.5 - La spéciation stasipatrique
Définition :
C'est une spéciation qui se produit au sein d'une espèce dont l'aire de répartition, très étendue, est subdivisée en plusieurs petites populations. Ces petites populations sont caractérisées par des réarrangements chromosomiques qui jouent un rôle majeur dans la spéciation en diminuant la viabilité ou la fécondité des hybrides.
Exemple : Les populations robertsoniennes de souris domestique (Mus musculus)
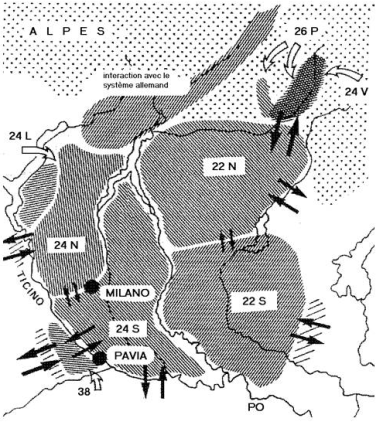
L'aire de répartition de chaque race est représentée en hachures sombres avec le nombre de chromosomes correspondant. Les hachures claires représentent les zones d'hybridation connues et les flèches noires les potentialités de flux génique fig. 2.15) (d'après Auffray 1988[1], avec l'autorisation de l'auteur)

Passage d'un nombre chromosomique de 2n = 40 à 2n = 22 (fig. 2.16) par l'intervention de 9 fusions de chromosomes acrocentriques donnant des chromosomes métacentriques (d'après Auffray 1988[1])
Dans la plaine du Pô (Italie), dans un rayon de 30 à 100 km autour de la ville de Milan, existent des populations chromosomiques de la Souris domestique caractérisées par une réduction du nombre des chromosomes par fusion robertsonienne. Il s'agit de populations liées à l'homme, peuplant des fermes de grosses exploitations agricoles et donc relativement isolées les unes des autres. On peut remarquer que ces populations à nombre réduit de chromosomes s'excluent géographiquement les unes les autres et ne cohabitent pas avec les populations « standards » à 40 chromosomes. Elles peuvent s'hybrider entre elles mais si les croisements entre les populations robertsoniennes et les populations à 40 chromosomes donnent des hybrides fertiles, les hybrides obtenus par croisement entre elles des populations à chromosomes fusionnés sont en général stériles par suite de difficultés d'appariement des chromosomes homologues lors de la métaphase de la division réductionnelle de la méiose.
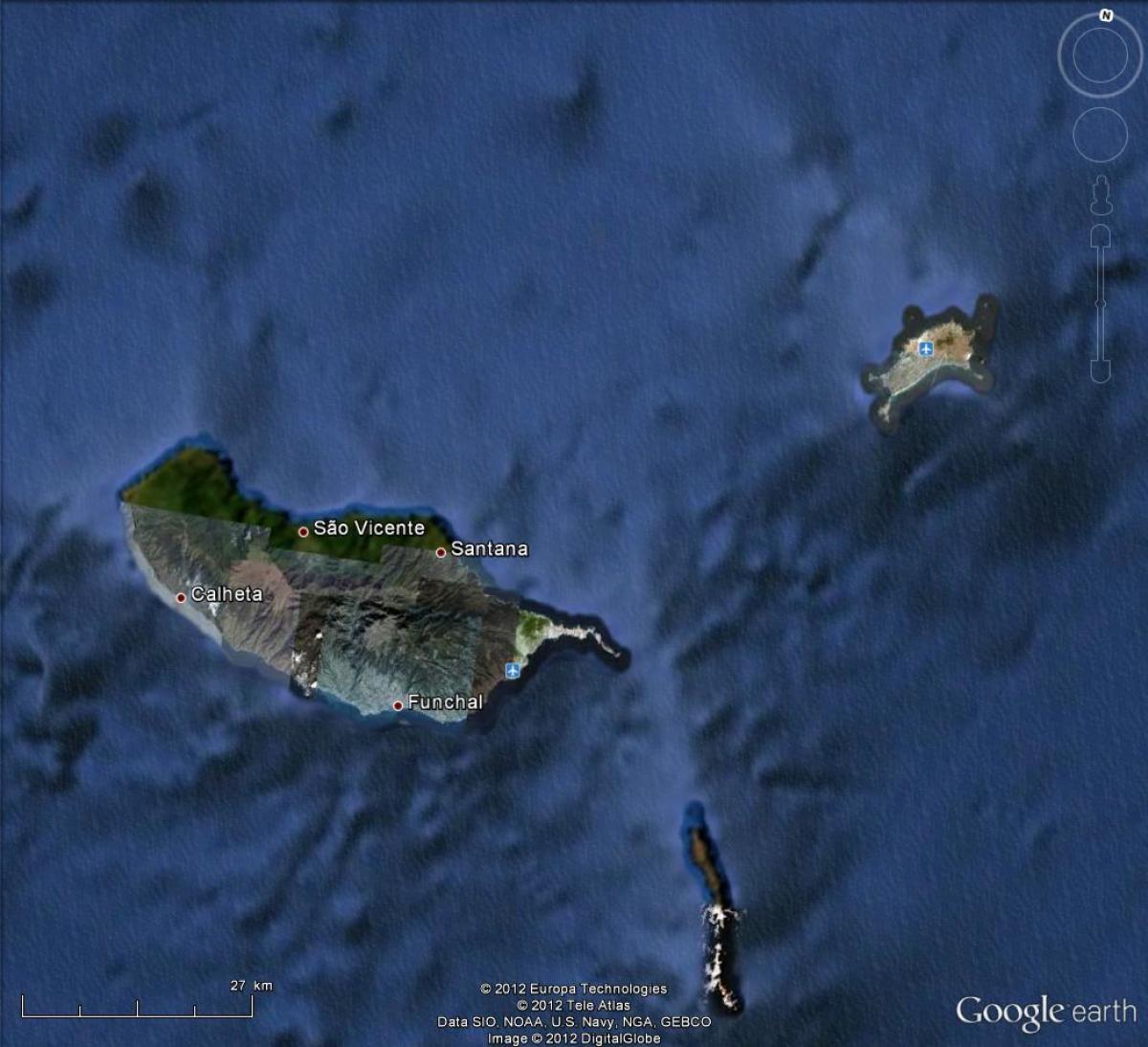
Récemment, des populations robertsoniennes de souris ont été découvertes sur l'Ile de Madère (fig. 2.17), avec six races chromosomiques. Parmi les 20 types de fusions robertsoniennes portées par les souris de Madère (nombre maximum de fusions dans une population = 9), 13 n'avaient jamais été observés jusque-là dans aucune population de souris et pour la première fois le chromosome 19 était concerné par de telles fusions. Cette radiation particulière, avec autant de populations sur une si faible surface, aurait été favorisée par la topographie de l'île, qui a isolé ces populations les unes des autres. Aucun hybride entre ces populations n'a été trouvé ce qui montre que les remaniements chromosomiques peuvent être un mécanisme efficace d'isolement reproducteur permettant une radiation sans être pour autant associé à un processus adaptatif. L'analyse génétique des allozymes a montré que ces souris sont plus proches des populations du Portugal que de toute autre région, mais leur ADN mitochondrial suggère pour sa part une origine plus nordique. Ces souris auraient donc été amenées sur l'île par les colonisateurs portugais à la fin du XVe siècle, mais un apport par l'intermédiaire des navigateurs vikings vers l'an 900 semble très probable. Les fusions seraient donc apparues avec une vitesse minimale de l'ordre de 9 en 500 à 1 000 ans. On peut noter que curieusement, toujours dans l'archipel de Madère, les populations de souris de l'île de Porto Santo ne présentent aucune fusion robertsonienne.
Ces populations semblent toutes dériver d'un nombre limité d'émigrants, que ce soit ceux qui ont colonisé les différentes fermes de la plaine du Pô, ou ceux qui ont été amenés par l'homme sur l'Ile de Madère. Ces petites populations sont caractérisées par des réarrangements chromosomiques qui jouent un rôle majeur dans la spéciation en diminuant la viabilité ou la fécondité des hybrides. Une fusion serait apparue une seule fois chez un individu dans chacune despetites populations isolées. Sous l'effet de la dérive méiotique et de la consanguinité le métacentrique passe à l'état homozygote. Plus l'effectif de la population est petit, plus la fixation est rapide. Les hybrides hétérozygotes étant moins fertiles du fait des problèmes d'appariements des homologues à la méiose, l'isolement des homozygotes devient plus marqué et les flux géniques avec la population d'origine se réduisent. L'adaptation aux conditions écologiques locales se met alors en place. Cela aboutit à l'occupation d'une nouvelle niche écologique et la nouvelle population peut coloniser l'aire de répartition de la population originelle. Ce processus est encore accentué par la fixation de nouvelles fusions. Néanmoins, si ce modèle est convaincant en ce qui concerne l'efficacité des barrières d'isolement reproducteur entre espèces à caryotypes remaniés, il reste de nombreuses zones d'ombre concernant l'apparition des fusions à l'état homozygote et leur diffusion dans les populations. Certains résultats suggèrent que ce phénomène puisse être particulièrement rapide, un délai d'une vingtaine d'années ayant été déterminé pour l'apparition et la fixation d'un échange de bras entre chromosomes fusionnés dans des populations robertsoniennes de souris de la plaine du Pô.