7.3 - Morphologie interne
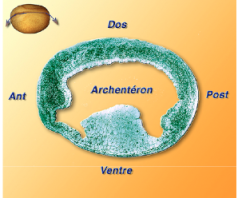
En morphologie interne, la neurula est creusée d'une large cavité archentérique. Une ouverture longitudinale de la région dorsale permet de rendre compte de cette cavité (Fig.191-video1).

L'archentéron sépare la fine région dorsale (avec le neuroderme et le mésoderme axial) de la région ventrale épaissie par l'endoderme végétatif Fig.192).
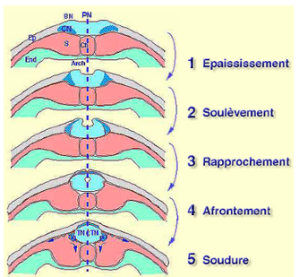
L'histologie permet de suivre l'évolution et la différenciation des feuillets dans la région dorsale. Le neuroderme s'enroule en passant de l'étape plaque neurale à celui de gouttière neurale. Après la soudure des bourrelets neuraux, le neuroderme forme un tube neural.
A la limite du neuroderme et de l'épiderme un territoire se distingue, c'est celui des crêtes neurales qui évolue différemment de l'épithélium du tube neural (Fig.193, Fig.194-video2).
Ainsi, le neuroderme se divise en deux catégories de tissus : l'épithélium neural à l'origine du nevraxe (moelle épinière et cerveau) et les crêtes neurales à l'origine de nombreux dérivés tels que les ganglions rachidiens et les chromatophores entre autre.

On s'aperçoit également que, parallèlement à la morphogenèse du tube neural, le mésoderme dorsal commence à scinder en ses principaux dérivés dorsaux. Ce sont, la chorde dorsale sur le plan médian et les somites latérodorsaux.
Au niveau cellulaire, l'épithélium neural est formé de cellules épithéliales hautes colomniformes dont certaines subissent une constriction apicale. De proche en proche, toutes les cellules effectuent la même transformation. Le résultat est un raccourcissement de la face externe de la plaque neurale qui génère une courbure concave. Ainsi se forme la gouttière neurale (Fig.194).
Au niveau subcellulaire, cette transformation est largement sous-tendue par le cytosquelette. L'axe baso-apical des cellules colomniformes est sous-tendu par des faisceaux de microtubules (Fig.195).
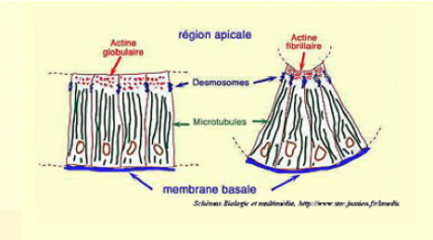
A la base, les cellules sont reliées par la membrane basale. A l'apex, les cellules sont solidaires grâce à des desmosomes et des jonctions sériées. Le cytoplasme apical est riche en actine globulaire qui se polymérise en actine fibrillaire. La contraction des fibrilles d'actine provoque la constriction apicale. Ce mouvement suffit à expliquer l'enroulement de la plaque neurale.
Comme dans le cas de la ségrégation des tissus blastuléens, la séparation entre le neuroderme et l'épiderme obéit à des différences d'adhérence et de reconnaissance cellulaire.
Sur la base des expériences de Holtfretter, il est possible de dissocier les tissus de la neurula (Fig.196).
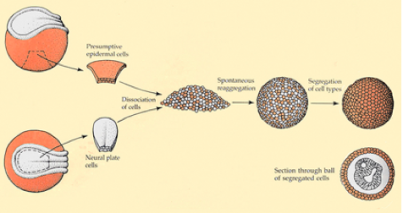
Des explants d'épiderme ventral et de neuroderme de la plaque neurale sont dissociés dans un milieu dépourvu de Ca++et de Mg++. A ce stade, il est nécessaire d'effectuer, en plus, une digestion enzymatique ménagée des protéines d'adhérence et des jonctions à l'aide de la trypsine. Les deux lots de cellules sont mélangés, les cellules adhèrent et se ségrégent en fonction de leur provenance. Finalement, après 3 jours de culture, le neuroderme se reconstitue entouré par l'épiderme.
Les cadhérines sont associées à ces mécanismes de reconnaissance cellulaire. La E-Cadhérine est associée à la formation des épithéliums embryonnaires, puis, au moment de la détermination du neuroderme, la E-Cadhérine se restreint à l'épiderme et la N-Cadhérine apparaît dans le neuroderme.
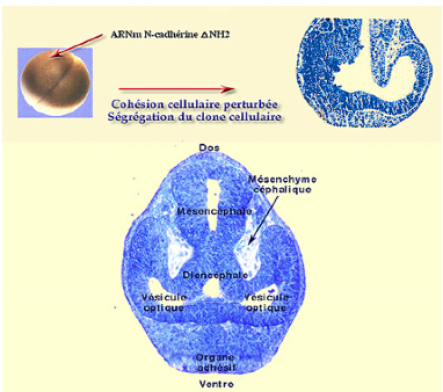
On peut montrer que la N-Cadhérine est indispensable à l'élaboration du neuroderme (Fig.197). L'injection d'ARNm de N-Cadhérine codant pour
une N-Cadhérine tronquée de son extrémité NH2 provoque des désordres et des perturbations de la cohésion cellulaire.