Vésicules tapissées de clathrine
A propos du Golgi nous avons déjà évoqué la clathrine, protéine impliquée dans le bourgeonnement de vésicules issues du réseau trans-golgien et la récupération de protéines à partir des endosomes précoces ou tardifs. De plus, comme la cavéoline, la clathrine participe aussi à l'endocytose en formant des vésicules de 100 à 150 nm de diamètre. Les vésicules d'endocytose tapissées de clathrine sont présentes dans toutes les cellules et en particulier sont impliquées dans la capture : de lipoprotéines chargées en cholestérol, de ferritine chargée en fer, de récepteurs occupés par des cytokines ou des hormones. Enfin, l'excès de membrane consécutif à l'exocytose est résorbé par ces vésicules, évènement qui recycle également la machinerie de fusion.
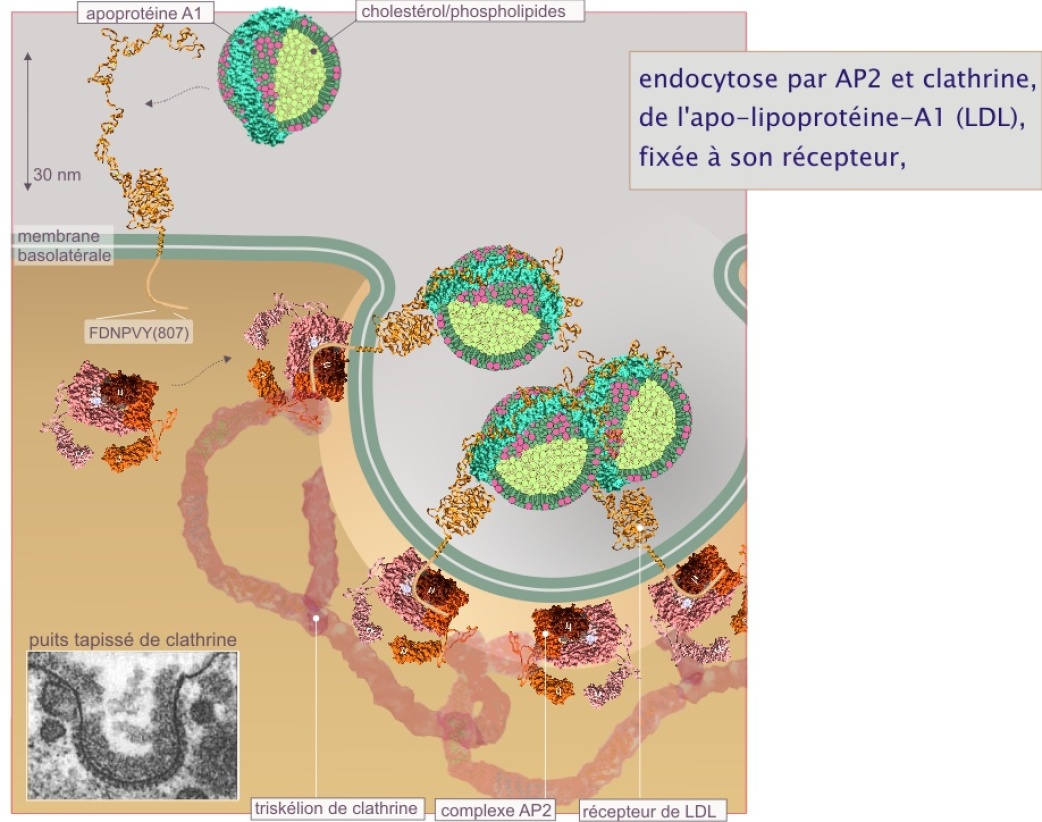
L'endocytose clathrine-dépendante commence par le recrutement de complexes AP2 (adapter protein–2) sur la membrane plasmique (l'évènement qui déclenche le recrutement semble être la production locale de l'inositol–4,5–diposphate ou inositol–3,4,5–triphosphate). AP2 est composé de quatre sous-unités, deux grandes dénommées \(\alpha\)– et \(\beta\)–adaptine (100 kDa), et deux petites, \(\mu\) et \(\sigma\) (50 et 20 kDa respectivement). AP2\(\mu\) est le récepteur du complexe (jouant un rôle équivalent à Sec23 dans la formation de la vésicule COPII, ou équivalent à AP1A\(\mu\) dans la formation du lysosome primaire) ; il interagit avec des peptides de destination porteurs du motif YXX\(\phi\)(dans lequel \(\phi\) représente l'un des acides aminés hydrophobes). Exemples : le récepteur de la transferrine est lié à AP2\(\mu\) grâce à la séquence YTRF et le récepteur de l'EGF grâce à la séquence YKGL. Plusieurs récepteurs membranaires différents peuvent partager la même vésicule tapissée de clathrine (voir figure 13 et pour plus de détail sur AP2 et le récepteur de LDL fixant l'apolipoprotéine–A1 ( = apoprotéine–A1 + cholestérol/phospholipides)).
Remarque :
Le récepteur à la lipoprotéine de faible densité (LDL) est reconnu par AP2 grâce à la séquence FDNPVY–807, qui est un motif canonique (consensuel). Ce motif fut découvert chez des patients atteints d'hypercholestérolémie (niveau très élevé de cholestérol sanguin) car déficients dans la capture de LDL par suite d'une substitution de la tyrosine 807 du motif d'internalisation.
L'auto assemblage du manteau AP2 provoque un regroupement de protéines membranaires dans les « puits tapissés », phénomène connu sous le nom de « patching » ou « capping ». Les complexes AP2 assemblés fixent alors la clathrine, complexe protéique hexamérique composé de trois chaînes lourdes (190 kDa) et de trois chaînes légères (LCa ou LCb de 30 kDa). L'ensemble forme une structure à trois «jambes» qui lui a valu le nom de triskélion (du Grec skelos = jambe). Les triskélions s'auto assemblent en hexagones et pentagones (voir figure 14) et finissent par former une structure grillagée formant une cage sphérique. C'est là l'origine du mot clathrine = cage

La fission qui s'ensuit nécessite l'intervention de la GTPase dynamine (protéine de 100 kDa), qui, en se liant au GTP, se polymérise pour former un col d'étranglement qui se resserre pour libérer la vésicule (un processus également dépendant de la présence d'inositol–4,5–disposphate). La dynamine recrute l'endophiline, phospholipase qui catalyse la production d'acide lysophosphatidique, qui autorise une courbure très forte de la membrane plasmique.
L'ensemble des évènements s'effectue en 60 secondes approximativement et on estime que, dans ce laps de temps, une cellule est capable de soustraire jusqu'à 40% de ses composants membranaires. Peu après son internalisation, la vésicule perd son manteau par l'intervention de Hsc70 (membre de la famille de chaperonnes « heat-shock »), d'auxiline et de synaptojanine (une phosphatidyl 5–kinase).
![]() Pour en savoir plus, consultez les documents suivants : « clathrine coat formation Wakelam » [pdf] (408 Ko), « AP2 clathrin minireview » [pdf] (98 Ko).
Pour en savoir plus, consultez les documents suivants : « clathrine coat formation Wakelam » [pdf] (408 Ko), « AP2 clathrin minireview » [pdf] (98 Ko).