Les lipides en double couche
Remarque : Lipides et protéines
Les cellules sont entourées par la membrane plasmique qui est essentiellement une barrière indispensable entre le cytoplasme et le milieu extra-cellulaire. La membrane plasmique est un film très fin constitué de molécules protéiques (50% de la masse) et lipidiques (également 50% ). Il faut bien comprendre que la taille des molécules lipidiques est petite (environ 700 Da) comparée à celle des molécules protéiques (> 10.000 Da). Dans la membrane il y a donc beaucoup plus de molécules lipidiques que de molécules protéiques.
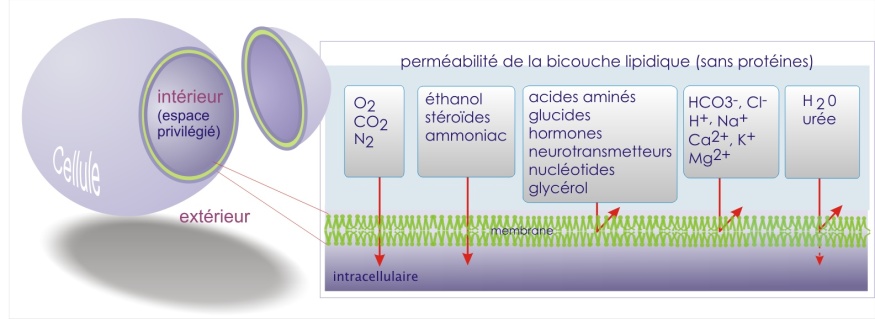
Les lipides forment une double couche (épaisse de 5 à 6 nm) qui est relativement imperméable au passage de la plupart des molécules hydrosolubles (protéines, hormones, ions) (voir figure 5). Cette propriété est bien illustrée par une expérience de diffusion des ions potassium ( K+ ). Alors que les ions K+ arrivent à un équilibre de diffusion dans l'eau sur une distance de 6 nm en 5 millisecondes, il leur faut 12 jours pour arriver à l'équilibre à travers une double couche de lipides. Même les petites molécules comme le glucose ou l'adrénaline mettent un temps considérable pour traverser. La membrane est donc une barrière très efficace ! Mais elle peut facilement être franchie par des molécules hydrophobes telles que les alcools, les stéroïdes et les anesthésiques généraux (pentobarbital).
Les propriétés de perméabilité d'une double couche lipidique artificielle sans protéines sont indiquées dans la figure 5 ci-dessous.
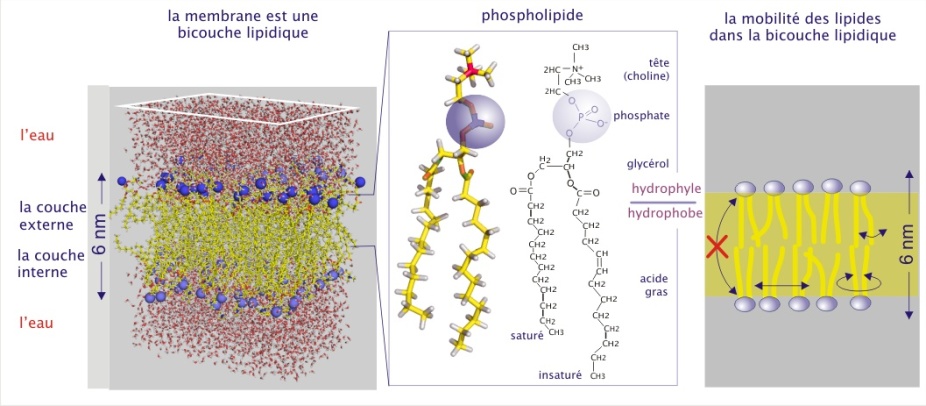
La structure en double couche est due aux propriétés amphiphiles des molécules lipidiques. Celles-ci possèdent ainsi une extrémité hydrophile (aimant l'eau ou polaire) et une extrémité hydrophobe (craignant l'eau ou apolaire) (voir figure 6).
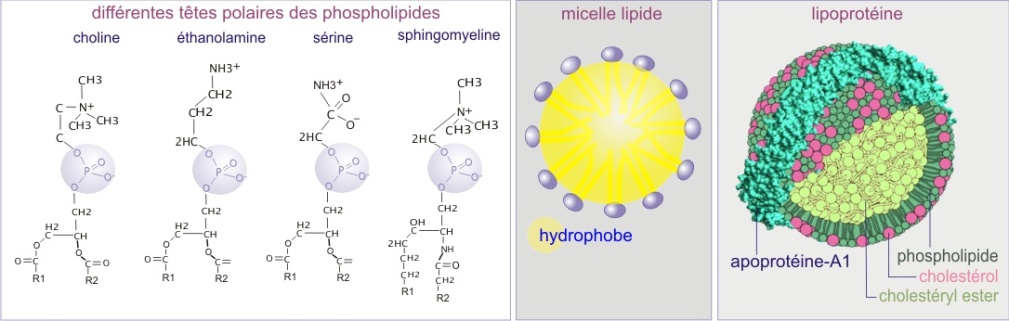
Il existe une grande variabilité de lipides membranaires. Les plus abondants sont les phospholipides qui sont composés d'une tête polaire contenant un groupement phosphate et de deux bras hydrocarbonés présentant ou non une certaine courbure (acides gras). Dans un environnement aqueux, les têtes polaires s'orientent vers l'extérieur et les bras apolaires vers l'intérieur de la membrane. La double couche lipidique est fluide du fait de la double mobilité, latérale et de rotation, des lipides (regardez l'animation ci-dessous "Configuration et mouvements des phospholipides").
Il y a peu d'échanges de lipides d'une couche à l'autre de la membrane (mouvements verticaux ou flip-flop), ce qui permet l'obtention de distributions asymétriques des différents lipides et confère ainsi des fonctions discriminatives aux feuillets membranaires selon qu'ils sont orientés vers l'extérieur ou vers le cytosol. Dans la membrane de l'hématie humaine, par exemple, tous les lipides contenant la choline, la phosphatidylcholine, la sphingomyéline et les glycolipides se trouvent à l'extérieur, alors que, pour la plupart, les phosphatidyléthanolamines et les phosphatidylsérines sont présents sur le feuillet interne. Dans un environnement aqueux, les lipides membranaire peuvent adopter deux autres configurations : la micelle au l'association avec une protéines (lipoprotéine) (voir figure 7).
![]()
Macromedia Flash - 49,7Ko
Les lipoprotéines jouent un rôle important dans le transport des lipides dans l'organisme mais sont aussi impliquées dans la distribution des lipides dans la cellule.
Macromedia Flash - 10,4Ko
Les lipides sont synthétisés dans le réticulum endoplasmique lisse et c'est là que l'asymétrie des couches est engendrée par des protéines d'échange des phospholipides (lipoprotéines intracellulaires). L'asymétrie lipidique est importante sur le plan fonctionnel surtout dans la localisation des protéines liées à la membrane et intervenant dans la transduction du signal.
Certains lipides sont glycosylés : les glycolipides (voir figure 8).
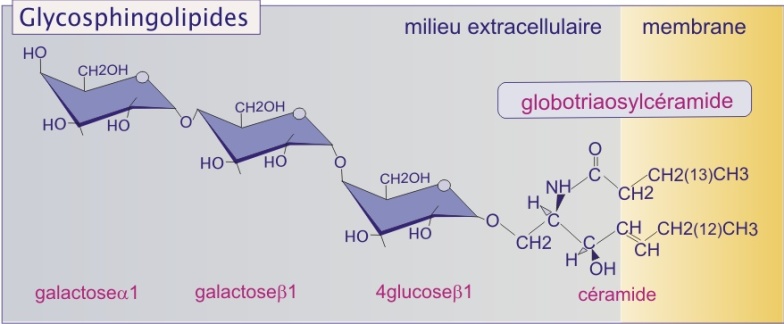
Les résidus glucidiques, tels que galactose, glucose mais aussi acide sialique, sont ajoutés dans l'appareil de Golgi. Les glycolipides sont toujours associés au feuillet membranaire externe et appartiennent à un ensemble que l'on appelle le glycocalyx. Le glycocalyx est la zone périphérique cellulaire riche en glucides (voir plus loin la page "Différentes protéines sont associées aux membranes de différentes façons"). Parce que les résidus glucidiques liés avec les protéines sont souvent impliqués dans les interactions de la cellule avec son environnement, il est possible que les glycolipides jouent un rôle analogue (cf. les sélectines dans la ressource sur les molécules d'adhérence).
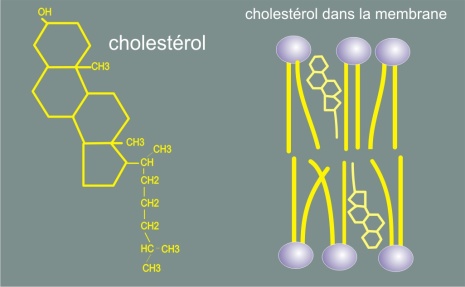
Le cholestérol est un lipide de structure distincte. Il joue un rôle particulier au sein de la membrane (voir figure 9), en la rendant moins déformable (plus rigide) et en diminuant sa perméabilité aux petites molécules hydrosolubles.
![]() Pour en savoir plus sur le modèle de la mosaïque fluide des membranes cellulaires par Singer et Nicolson en 1972 :
Pour en savoir plus sur le modèle de la mosaïque fluide des membranes cellulaires par Singer et Nicolson en 1972 :
ci-joint un article de revue historique dans lequel notre compréhension actuelle de la composition et de l'organisation de la mambrane cellulaire a été révélé. Les auteurs Singer et Nicolson, proposent le "modèle de la mosaïque fluide des membranes cellulaires [pdf]", où les lipides forment une bicouche dans laquelle les protéines sont insérées de différentes manières.