Acides nucléiques : ADN et ARN
ADN et ARN sont des polymères de nucléotides. Un nucléotide comporte une base azotée (composée d'un ou deux cycles carbonés), un (désoxy)ribose (glucide pentose cyclique) et un ou plusieurs groupements phosphates. Le polymère s'assemble par liaison phosphodiester entre les riboses sur les carbones 5t et 3t. Les riboses-phosphates forment le squelette du polymère souvent présenté sous forme de ruban (voir figure 2a). Les chaînes nucléotidiques ont une grande souplesse conformationnelle, ce qui autorise l'apparition de structures complexes. Ces structures (tertiaires) sont dues à l'appariement spécifique des bases par des ponts hydrogènes, trois entre C et G et deux entre A et T ou A et U, mais aussi à la tendance qu'ont les bases à s'empiler les unes sur les autres (« Pi stacking ») grâce aux charges électrostatiques et aux forces de Van der Waals (voir figure 2a).
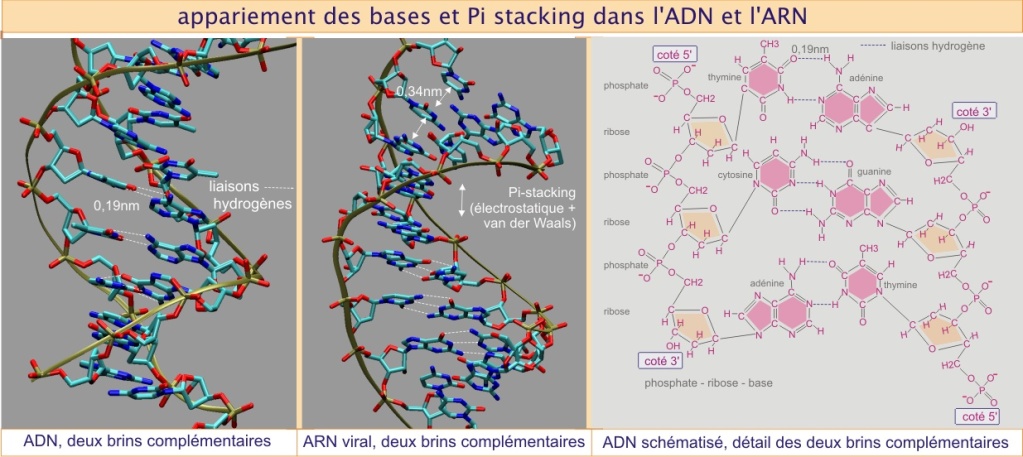
Rappel : Structure de l'ADN
C'est dans leur célébrissime publication parue dans Nature en 1953, que Watson et Crick élucident la structure de l'acide désoxyribonucléique en proposant le modèle moléculaire de deux chaînes hélicoïdales enroulées autour d'un même axe (« double hélice »). L'originalité de cette structure réside dans la manière dont les deux chaînes parallèles sont solidarisées par l'association d'une base purique à une base pyrimidique sur le même plan perpendiculaire à l'axe de la chaîne. Les bases établissent entre elles des liaisons hydrogènes obligées. En effet, selon le modèle proposé qui s'avéra exact, l'adénine (purine) ne peut se lier qu'à la thymine (pyrimidine) et la guanine (purine) qu'à la cytosine (pyrimidine) : c'est le principe de « » typique de l'ADN.
Dans le cas de l'ADN, deux chaînes nucléotidiques complémentaires permettent la réalisation de toutes ces interactions, si bien qu'une double hélice très stable apparaît (appariement intermoléculaire). Dans le cas de l'ARN cette complémentarité entre les chaînes est rare, mais possible comme cela est montré sur le panneau central de la figure 2a, qui concerne l'ARN viral. En général, chez les eucaryotes, la structure de l'ARN résulte des appariements intramoléculaires. Voir par exemple les ponts hydrogènes présents dans l'ARN de transfert (indiqués par des lignes pointillées dans la et qui sont à l'origine de la conformation complexe présentée en .