Ep. 4 - Principales caractéristiques fongiques (2/2)
Modalités de reproduction
Les mycètes présentent également des particularités au niveau de leurs modalités de reproduction. D'une manière générale, on considère que les champignons peuvent se reproduire de 2 manières : de façon sexuée et de façon asexuée. La reproduction qu'elle soit sexuée ou asexuée va aboutir à la formation de structures fongiques appelées « spores ». Ces spores sont formées à partir d'un pied-mère, vont être capables de s'en détacher et, dans des conditions environnementales favorables, de germer pour développer un thalle végétatif filamenteux. Ces spores assurent la conservation et la dispersion du mycète.
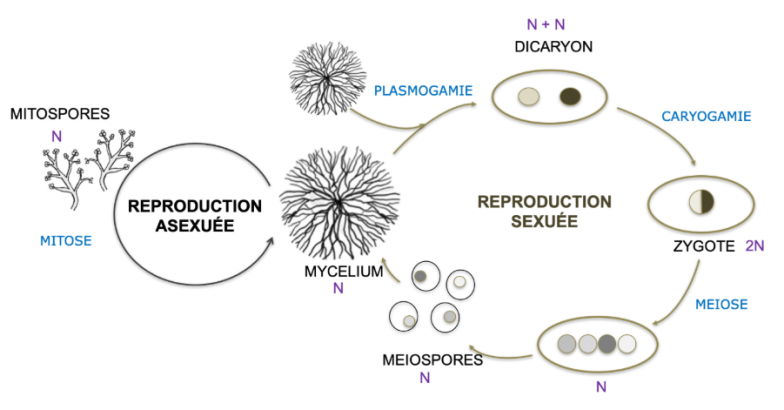
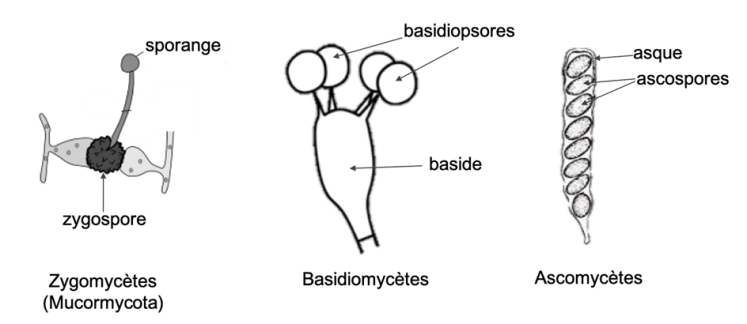
La reproduction sexuée implique une étape de fécondation entre des cellules de 2 individus, ce qui aboutit à la formation d'un zygote diploïde. Ce zygote va subir la méiose et donner naissance à 4 cellules haploides qu'on appelle des méiospores ou spores sexuées. Les méiospores chez les Ascomycètes sont appelées des ascospores et sont généralement regroupées par 8 au sein d'une cellule particulière appelée asque. Comme on le verra plus tard, ces asques peuvent être régroupés au sein de structures particulières. Chez les basidiomycètes, la méiose a lieu au sein de cellules appelées basides, situées généralement dans l'hymenium au sein des chapeau du sporophore. Les 4 noyaux haploïdes ainsi formés sont incorporés chacun dans une spores sexuées appelées basidiospores qui se forment sur de minuscules tiges à l'extérieur de la baside. Chez les Zygomycètes, des hyphes compatibles, contenant chacun de multiples noyaux haploïdes, vont fusionner. Les noyaux haploïdes vont également fusionner 2 à 2 pour donner une cellule appelée Zygospore contenant donc de multiples noyaux diploïdes. Les noyaux à l'intérieur de la zygospore subissent ensuite une méiose et simultanément un sporange se forme à partir de la zygospore. Chaque sporange contient les produits de la méiose : des noyaux haploïdes contenus dans des spores. Ces spores se dispersent et germent pour former une nouvelle génération d'hyphes haploïdes.
La reproduction asexuée est aussi appelée « multiplication végétative », Elle ne concerne qu'un individu isolé et n'implique pas de fécondation. Il s'agit simplement de la différenciation de spores par simples mitoses. Les cellules de la colonie portent toutes le même contenu génétique mais les spores asexuées ou mitospores présentent une morphologie particulière. Elles vont en outre pouvoir se détacher de la colonie d'origine et germer sous de bonnes conditions. Les champignons en quelque sorte maîtrisent le clonage haut débit car ces mitospores sont bien des clones et leur production est souvent massive. Les spores asexuées présentent des morphologies très variées. Chez les champignons inférieurs, leur production est de type endogène, à l'intérieur d'un réceptacle que l'on peut appeler mitosporange. Chez les chytridiomycètes, les mitospores sont appelées zoospores et sont monoflagellées. Chez les Zygomycètes, les mitosporanges renferment des mitospores non flagellées. Les mitospores qui présentent le plus de variabilités sont observées au sein des ascomycètes, on les appelle aussi conidies. Leur production peut être endogène ou exogène (dans ce cas directement à l'air libre). Les mitospores ou conidies chez les Ascomycètes peuvent être uni- ou pluricellulaires, de forme variable, pigmentées ou non... Chez les Basidiomycètes, de façon étonnante, peu d'espèces sont capables de produire des mitospores.
Les spores (sexuées ou asexuées) peuvent être considérés comme des organes de dissémination et/ou conservation. Elles sont dispersées par le vent, l'eau, des animaux (vecteurs) et peuvent rester viables pendant de longues périodes et surmonter des conditions de vie défavorables (dessiccation, carences, gel...). Elles sont présentes partout et particulièrement dans l'air.
Structure pariétale
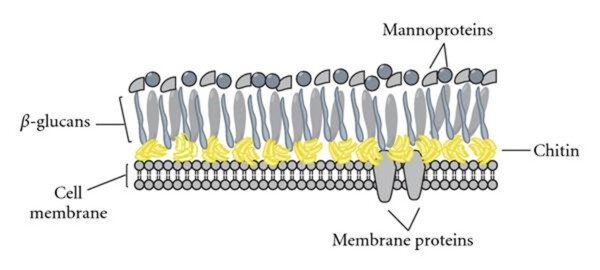
Une autre particularité concerne la paroi fongique (structure située autour de la membrane plasmique) qui possède une composition spécifique chez les mycètes. Cette paroi est particulièrement importante pour le fonctionnement cellulaire. Tout d'abord, elle assure un soutien structurel et règle la forme de la cellule. Sans paroi, les cellules sont sphériques et la différentiation de filaments n'est plus possible. Ensuite, elle joue un rôle de protection contre les facteurs environnements (froid, dessiccation, stress mécanique ou osmotique...) mais aussi contre d'autres micro-organismes. Elle participe d'autre part aux mécanismes d'adhésion cruciaux pour la colonisation de certains environnements et la formation de biofilms fongiques. Pour les champignons pathogènes, la paroi cellulaire est essentielle pour la virulence et la pathogénicité. Elle offre à la fois des propriétés adhésives essentielles pour l'invasion des tissus de l'hôte et une protection contre les mécanismes de défense de l'hôte. Globalement, on peut dire que les parois fongiques sont constituées d'éléments communs notamment : chitine/chitosane, des glucanes et des glycoprotéines. Néanmoins, on constate une variabilité importante dans la composition et l'organisation des parois selon les espèces mais aussi selon le type d'organes (spores ou hyphes par exemple). Chez la plupart des espèces fongiques, la couche interne de la paroi est relativement conservée et est constituée d'un ensemble de glucanes ramifié et de chitine. Le glucane et la chitine forment des liaisons et s'assemblent en microfibrilles qui constituent un exosquelette qui résiste notamment à la pression interne substantielle exercée sur la paroi par le cytoplasme et la membrane. À cette structure centrale solide et résistante sont ajoutés une variété de protéines et d'autres composants superficiels, qui forment la couche externe , dont la composition peut varier en fonction de l'espèce fongique.
La chitine est un élément bien connu des parois fongiques. Il s'agit d'un polymère de N-acétylglucosamine, que l'on retrouve également comme élément constitutif des exosquelettes d'insectes, mollusques et crustacés. La chitine représente entre 1 et 15 % de la masse de la paroi cellulaire, les mycètes de type levures ayant généralement 1 à 2 % de chitine et les champignons filamenteux étant plus riches jusqu'à 15 % de chitine. Ce composé est synthétisé par des enzymes appelées chitine synthases associées à la membrane plasmique et elle va être extrudée dans l'espace de la paroi cellulaire sous forme de polymère linéaire. Cette chitine peut être modifiée par l'action de chitine deacetylases pour générer du chitosane, un polymère de chitine déacétylée. Le chitosane est beaucoup plus soluble que la chitine et on en sait beaucoup moins sur son rôle exact. La quantité de chitine qui est convertie en chitosane semble différente selon les espèces de champignons et peut également varier selon le type de cellule. Il apparaît par exemple que les parois de nombreux zygomycètes sont particulièrement riches en ce composé.
Les glucanes sont des polymères de glucose qui diffèrent entre eux par la nature de la liaison entre glucose. Le β-(1,3)-Glucane est un constituant majeur de toutes les parois cellulaires fongiques représentant entre 30 et 80 % de la masse de la paroi. D'autres types de glucanes, comme le β-1,6-glucane et l'𝛼-1,3-glucane, peuvent aussi être inclus. Ces glucanes sont synthétisés par des glucane-synthases associées à la membrane plasmique, qui libèrent des polymères de glucane linéaires nouvellement formés dans l'espace de la paroi cellulaire.
Un certain nombre de glycoprotéines ont été trouvées associées à la paroi cellulaire. Ces protéines contiennent des oligosaccharides ainsi que des polymères riches en mannose : des mannanes et galactomannanes. Certaines glycoprotéines sont liées de manière covalente à la matrice de la paroi cellulaire. Le glycosylphosphatidylinositol ou GPI est un glycolipide qui va par exemple permette l'ancrage de certaines de ces protéines à la membrane plasmique. Les rôles de ces protéines sont variés: de nombreuses ont pour action de lier ensemble les constituants pariétaux et créer ainsi la matrice tridimensionnelle de la paroi, elles interviennent aussi pour remodeler la paroi cellulaire afin de permettre la ramification et l'anastomose. Elles sont donc cruciales pour la biogenèse de la paroi cellulaire. D'autres fonctionnent comme des protéines senseurs détectant les modifications externes, comme des adhésines qui interagissent avec d'autres cellules, ou sont impliquées dans l'acquisition du fer et dans la détoxication par radicaux libres de l'oxygène.
D'autres constituants pariétaux sont optionnels selon les espèces et organes considérés. Les mélanines par exemple sont des polymères formés de composés phénoliques et sont considérés comme des pigments favorisant la survie des cellules. On les trouve fréquemment dans les parois des conidies, des ascospores et d'autres structures de conservation (les sclérotes par exemple). On sait qu'il existe au moins deux voies de synthèse des mélanines chez les mycètes, chaque mycète utilisant généralement une seule de ces 2 voies.
Les hydrophobines sont des petites protéines qui sont sécrétées par certains champignons et se positionnent au niveau de la superficie des parois. Elles forment une classe de protéines avec des séquences protéiques globalement assez peu conservées à l'exception de cystéines régulièrement espacés. Ces cystéines peuvent établir des liaisons covalentes (ponts disulfures) permettant de générer une structure tertiaire avec des propriété amphiphile, c'est-à-dire une protéine avec un pôle hydrophobe et un pôle hydrophile. Une autre de leur caractéristique est que ces protéines peuvent s'auto-assembler pour former des rodlets, générant une interface hydrophobe entre les champignons filamenteux et leur environnement. Les rodlets forment une monocouche autour de structures aériennes telles que les hyphes et les fructifications, recouvrant des surfaces hydrophiles pour les rendre hydrophobes. Les hydrophobines jouent un rôle dans la morphogenèse et sont également impliqués dans l'adhésion des cellules fongiques aux surfaces et ont donc été associés à la virulence des champignons pathogènes des plantes et des insectes.
Autres spécificités fongiques
Depuis Saccharomyces cerevisiae en 1996, plus de 1500 génomes fongiques ont été séquencés. C'est plus que le nombre de séquences génômique disponibles chez les plantes (<400) et les animaux (<900). Les génomes des champignons intéressent depuis longtemps les généticiens parce que de nombreuses espèces sont rapides et faciles à cultiver, que leur ADN peut être facilement manipulé et que nombre de leurs processus cellulaires essentiels sont les mêmes que ceux de tous les eucaryotes, y compris les humains. L'une des caractéristiques génomiques qui rendent les champignons particulièrement aptes au séquençage du génome entier est la petite taille de leur génome (c'est-à-dire la quantité totale d'ADN qui compose un génome). Par exemple, la taille moyenne du génome des champignons n'est que de 44 Mb (mégabases), contre environ 3 400 Mb pour les mammifères et environ 5 020 Mb pour les plantes à fleurs. Les champignons sont également remarquables car ils comprennent l'espèce dont le génome est le plus petit jamais signalé pour un eucaryote. Cette espèce est le parasite intracellulaire obligatoire Encephalitozoon intestinalis, dont le génome ne comprend que 2,3 Mb d'ADN - c'est-à-dire plus de 1000x plus petit que le génome humain. A l'opposé, le plus grand génome fongique estimé présente une taille de 5 800 Mb chez Neottiella, un ascomycète qui parasite les mousses.
Une autre particularité fongique qui mérite d'être rapportée ici est la présence d'ergostérol membranaire. C'est le stérol le plus abondant dans les membranes des cellules fongiques, où il régule la perméabilité et la fluidité de celles-ci. En raison de ses fonctions cruciales associé au fait que l'ergostérol est présent dans les membranes cellulaires des champignons, mais absent dans celles des animaux (où il est remplacé par le cholestérol), la biosynthèse de l'ergostérol reste la cible d'un grand nombre d'antifongiques et fongicides. À titre d'exemple, l'amphotéricine B, un médicament antifongique, se lie physiquement à l'ergostérol à l'intérieur de la membrane, créant ainsi un pore polaire dans les membranes fongiques. Dans le domaine de la protection des plantes de nombreux fongicides ciblent aussi l'ergostérol, mais agissent de manière différente, en inhibant des enzymes de la voie de biosynthèse de l'ergostérol.