

3.1-La Maturation
La progestérone se lie à un récepteur cytoplasmique, ce qui déclenche une cascade d'événements intra cytoplasmiques. En l'absence de progestérone, le récepteur cytoplasmique est complexé à des protéines HSP[1] qui le rendent inactif. En présence de progestérone, le récepteur s'associe à son ligand en se dissociant des HSP[1].
L'ensemble Progestérone plus récepteur entre dans le noyau, se dimérise et active la transcription des gènes d'intérêt. Le dimère se fixe sur des segments d'ADN, les Progestérone Responsive Elements .
La fixation du dimère progestérone-récepteur permet le recrutement d'activateurs de la transcription, qui se traduit par :
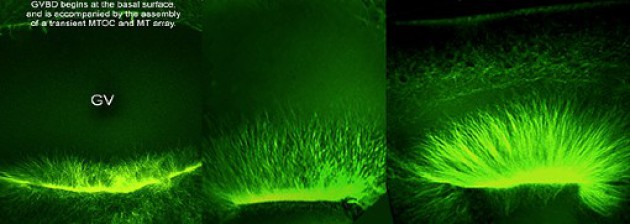
Dans un premier temps, on observe la formation d'un disque de cytosquelette riche en microtubules et en actine fibrillaire à la base de la vésicule germinative .
Puis les microtubules traversent la membrane nucléaire et entrent dans le nucléoplasme.
Les conséquences sont triples :
La membrane nucléaire se rompt à partir de la base de la VG[2].
Le réseau de microtubules ainsi formé coïncide avec la remontée de la VG[2] vers le pôle animal.
Enfin, les chromosomes en condensation sont transportés par les microtubules jusqu'au pôle animal. La première mitose se poursuit avec la métaphase, l'anaphase, la télophase puis l'émission au pôle animal du premier globule polaire.
Dans le détail, les événements s'effectuent de la manière suivante :
Première division réductionnelle
A la suite de la compaction, les chromosomes se polarisent sur le fuseau mitotique en formation. A ce stade, l'axe du fuseau est parallèle à la membrane plasmique. Au cours de la période d'élongation du fuseau, on observe la rotation de 90° de l'axe mitotique. Environ une heure à une heure vingt plus tard, le fuseau se retrouve perpendiculaire à la membrane plasmique. C'est à ce moment que débute la montée anaphasique des chromosomes. Le premier globule polaire est émis environ une heure et demie à deux heures après la reprise de la meiose. La première division réductionnelle est achevée et suivie immédiatement par la deuxième division équationnelle .
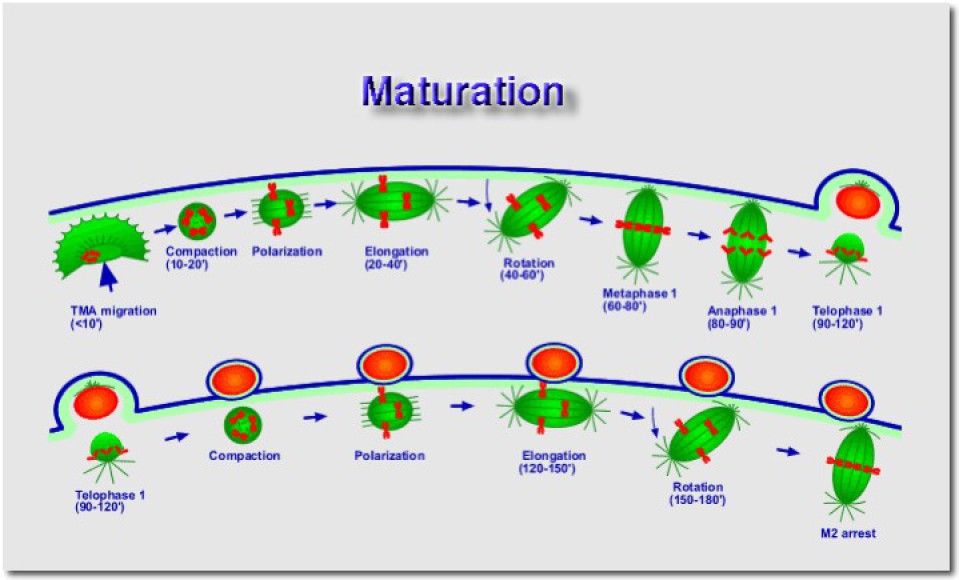
Deuxième division équationnelle
Après les phases de compaction, polarisation, élongation puis rotation, le fuseau mitotique se bloque en métaphase. L'ovocyte secondaire reste ainsi bloqué en métaphase de deuxième division de méiose (ou de maturation). Contrairement à l'ovocyte primaire, il est fécondable et c'est d'ailleurs la fusion avec le spermatozoïde qui déclenchera la reprise et l'achèvement de la méiose avec l'émission du deuxième globule polaire.
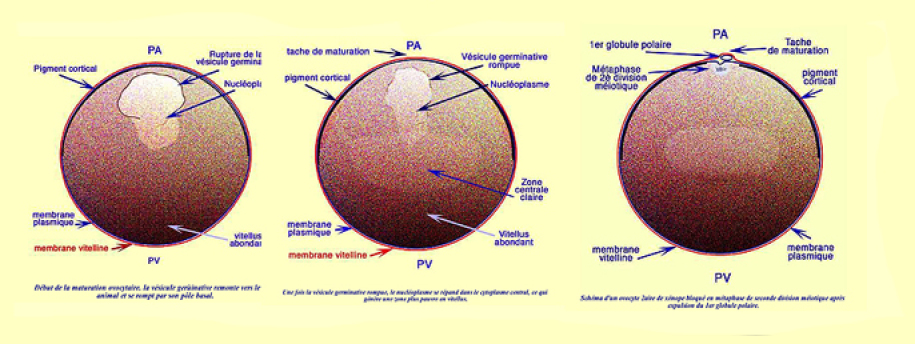
En morphologie externe, la remontée de la VG[2] laisse une trace plus claire que l'on appelle la tache de maturation. Au centre de cette tache plus claire se distingue un point sombre qui marque l'emplacement de la métaphase de deuxième division méïotique.
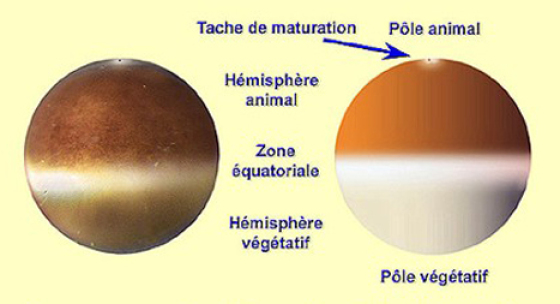
Au total l'ovocyte secondaire est toujours anisotrope et présente deux gradients inverses : le gradient vitellin croissant du PA vers le PVg hérité de l'ovogenèse, et puis un deuxième qui apparaît après la maturation : le gradient ribosomique inverse du précédent. En effet le contenu de la VG[2] s'est déversé dans le cytoplasme enrichissant la région animale en ARN ribosomiques .

Au niveau moléculaire la question qui se pose est de savoir comment la progestérone interrompt le blocage de l'ovocyte primaire.
Depuis longtemps, on suspecte l'existence d'un facteur de maturation : le MPF pour Maturation Promoting Factor. En réalité, ce facteur possède un rôle cyclique déjà connu dans la mitose. On l'appelle maintenant MPF, Mitosis Promoting Factor.
Le MPF[3] est formé de 2 sous-unités : la cycline B et la protéine P34. La P34 est une kinase dépendante de la cycline.
Les deux composants du MPF[3] sont déjà présents dans le cytoplasme de l'ovocyte mais inactifs. La progestérone convertit le complexe pré MPF[3] en MPF[3] actif. Plus précisément, la progestérone active la traduction de la protéine C-mos, une sérine-thréonine kinase, à partir d'ARNm stockés dans le cytoplasme.
Il s'en suit une cascade de phosphorylations qui se termine par l'activation de la protéine P 34. Le MPF[3] est alors activé. La protéine C-mos n'est donc d[3]étectable qu'à partir de la maturation de l'ovocyte. Le rôle crucial de C-mos est attesté par des expériences d'inhibition fonctionnelle utilisant des ARN antisens injectés dans le cytoplasme de l'ovocyte primaire. Dans ce cas, la rupture de la VG[2] et la maturation n'ont pas lieu.
Comment s'effectue le blocage en métaphase de deuxième division de maturation ?
Dès que la maturation est achevée, apparaît une protéine qui se combine avec C-mos, la protéine cdk2, une autre kinase cycline dépendante. Ces deux sous unités C-mos et cdk2 forment un complexe appelé CytoStatic Factor. Le MPF[3] devient alors inactif.
Le CSF[4] est connu dans la mitose pour bloquer la métaphase. On retrouve ainsi le CSF[4] dans le cytoplasme de l'ovocyte au moment où il se bloque en métaphase de deuxième division de maturation.
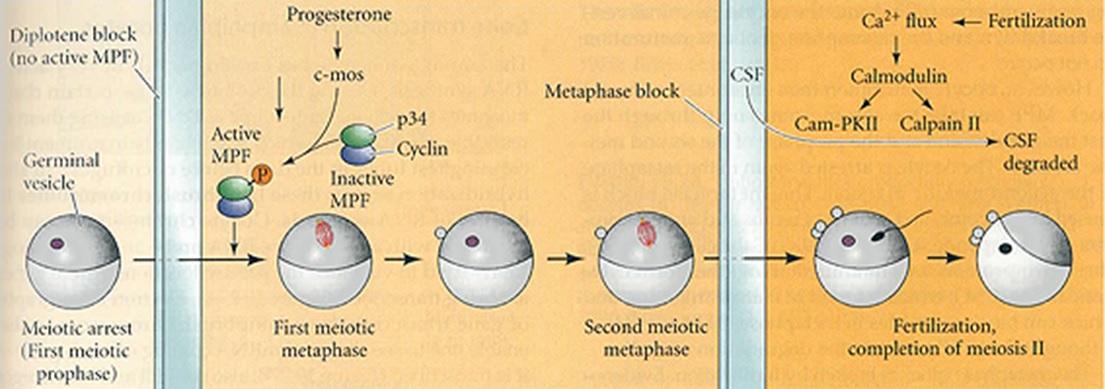
Le CSF[4] est dégradé au moment de la fécondation. Le blocage métaphasique est ainsi levé et la méiose s'achève par l'émission du deuxième globule polaire.
Il existe depuis longtemps des preuves expérimentales de la présence d'un facteur de maturation dans le cytoplasme de l'ovocyte en maturation
On sait cultiver « in vitro » des ovocytes de xénope. Si on ajoute de la progestérone dans le milieu de culture, on provoque artificiellement la maturation de tous les ovocytes au stade VI (fin de croissance).
L'expérience consiste alors à prélever par micro-ponction, du cytoplasme d'ovocyte en maturation et de le transférer par micro-injection dans des ovocytes primaires non maturés.
Les résultats montrent que les ovocytes micro-injectés par le cytoplasme en maturation entrent à leur tour en maturation.
Ainsi, après la maturation nous disposons d'un ovocyte secondaire fécondable bloqué en métaphase de deuxième division de maturation. Le déblocage se fera au moment de la fécondation, car le spermatozoïde va déclencher toute une série d'événements membranaires et cytoplasmiques qui donnent le départ du développement embryonnaire.




