Les caroténoïdes
Les caroténoïdes constituent la seconde famille de pigments photosynthétiques. Elle comprend plus de 600 membres. La structure chimique de ces pigments est variable et plusieurs volumes seraient nécessaires pour les décrire complètement. Dans cette ressource, vous allez découvrir les caractéristiques principales de cette famille de pigments.
La famille de caroténoïdes est divisée en deux sous-familles appelées les carotènes et les xanthophylles. Les carotènes sont des caroténoïdes ne contenant aucun atome d'oxygène.
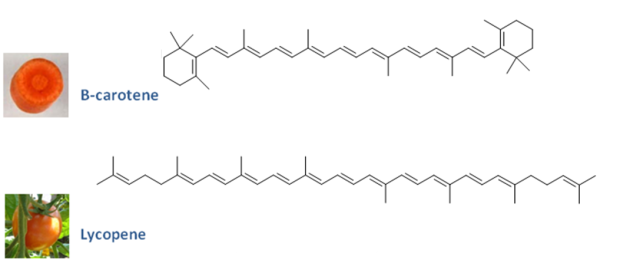
Les carotènes les plus couramment rencontrés sont le β-carotène et le lycopène, les pigments qui respectivement confèrent la couleur orangée des racines des carottes et la couleur rouge des tomates mûres (Figure 16). A première vue, ces deux carotènes ne se ressemblent pas du point de vue chimique. Ils possèdent cependant le même nombre d'atomes de carbone,40, et un réseau de doubles liaisons conjuguées.
Contrairement aux carotènes, les xanthophylles contiennent de l'oxygène. Celui-ci peut être présent sous différentes formes. Les principales sont de type époxy, hydroxy, cétone, hydrate de carbone, carboxyle, éther (Figure 17.)
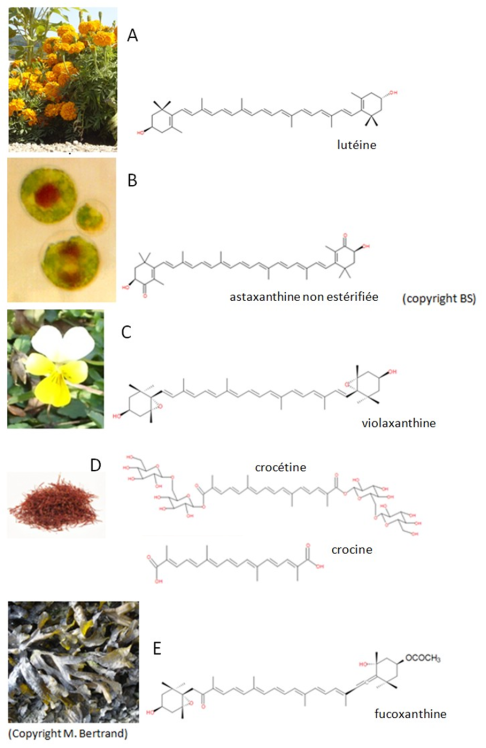
A. Les pétales des tagettes doivent leur couleur orange à l'accumulation d'une xanthophylle portant deux groupements hydroxyle, la lutéine.
B. En réponse à un stress, la microalgue rouge d'eau douce Haematococcus pluvialis synthétise de l'astaxanthine, une xanthophylle rouge caractérisée par deux groupements hydroxyles et cétones.
C. Certaines espèces de pensées (Viola sp.) accumulent dans leurs pétales une époxy-xanthophylle, la violaxanthine.
D. Le safran doit sa couleur à la crocétine et la crocine, deux xanthophylles portant respectivement des groupements carboxyl et hydrate de carbone. Ces deux fonctions permettent à ces deux pigments d'être solubles dans les solvants aqueux, ce qui est rare pour les caroténoïdes.
E. La fucoxanthine est probablement le caroténoïde le plus abondant à la surface du globe terrestre. Il est abondant chez les algues brunes (macroalgues les plus abondantes en terme de biomasse) et les diatomées (microalgues dominantes). Il se caractérise par la présence des différents groupements oxygénés présentés plus haut, auxquels s'ajoute une liaison éther et un carbone allénique (carbone lié à deux atomes de carbone par deux doubles liaisons).
Le spectre d'absorbance des caroténoïdes présente généralement trois bandes d'intensité inégale (Figure 18A). Ces bandes sont essentiellement présentes sur l'intervalle 400-500 nm. Cette propriété trouve son origine dans les doubles liaisons conjuguées des pigments et elle est en lien avec la couleur jaune-orangé des caroténoïdes. La position exacte des maxima d'absorption peut varier d'un caroténoïde à l'autre et aussi en fonction de l'environnement du pigment. Pour s'en convaincre, observons un exemple très spectaculaire dans le monde animal : le homard. Comme vous l'avez certainement déjà observé vous-même, la couleur du homard cuit est rouge-orangé (Figure 18B). L'analyse chimique révèle que la coloration est due à la présence dans l'exosquelette d'un caroténoïde rouge, l'astaxanthine. Alors comment se fait-il que l'exosquelette du homard vivant apparaisse bleu, comme vous pouvez l'observer dans la figure 18C ? Et bien, c'est simple. Dans la carapace, l'astaxanthine ne se trouve pas sous sa forme libre mais elle est attachée à une protéine. On parle de complexes protéine-pigment. Dans ces complexes, la molécule d'astaxanthine est “tordue” et le réseau de doubles liaisons conjuguées modifié de telle sorte que l'ensemble nous apparaisse bleu. Les parties plus sombres de l'exosquelette sont dues à une forte accumulation de complexes protéine-pigment. Elles sont en réalité bleu très foncé. Lors de la cuisson, la chaleur dénature les liaisons entre la protéine et l'astaxanthine, ce qui permet au pigment de reprendre sa configuration libre et d'apparaître rouge.
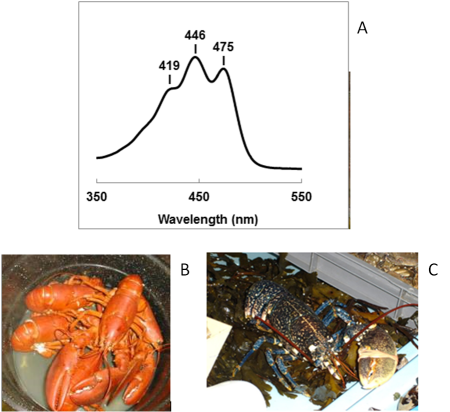
A. Spectre d'absorbance de la lutéine purifiée par chromatographie sur couche mince et solubilisée dans le méthanol pur. La structure en trois bandes du spectre d'absorbance de cette xanthophylle est bien visible. La position des maxima peut varier d'un caroténoïde à un autre et également d'un solvant à un autre.
B. Photographie de homards cuits. La couleur rouge vif de l'exosquelette est due à l'astaxanthine libérée des complexes protéiques au cours de la cuisson.
C. Photographie d'un homard vivant. La couleur bleue très foncée de l'exosquelette est due à l'accumulation de complexes protéine-astaxanthine. Dans ces complexes, le réseau de doubles liaisons conjuguées est « tordu » de telle sorte que son spectre d'absorbance est déplacé vers les grandes longueurs d'onde.