La nécessité de la structuration des molécules de pigments
Vous avez découvert dans la section "L'éclairement naturel et la mesure de son intensité". que l'activité photosynthétique pouvait être mesurée au travers de la mesure de la quantité d'oxygène émis au cours d'une illumination. Par ailleurs, vous vous rappelez les résultats des expériences d'Ingenhousz et Engelmann, à savoir que cette émission d'oxygène ne se déroule que dans les parties vertes des végétaux, c'est-à-dire celles qui contiennent de la chlorophylle. La présence de ces molécules constitue-t-elle une condition suffisante ? Pour le vérifier, illuminons une solution éthanolique de chlorophylle et mesurons la quantité d'oxygène émise (Figure 22).

Aucune émission n'est enregistrée. Au contraire une consommation d'oxygène est observée car la présence de lumière induit l'oxydation des molécules de chlorophylle, ce qui les dégrade. C'est le phénomène de blanchiment. A l'issue de l'illumination, la solution de chlorophylle est nettement moins verte qu'au départ (Figure 22). La présence des molécules de chlorophylle constitue donc une condition nécessaire à l'émission d'oxygène mais elle n'est pas suffisante. Nous pouvons donc faire l'hypothèse que pour que le processus photosynthétique fonctionne, il est indispensable que les molécules de chlorophylles soient dans leur état naturel. Que cela signifie-t-il ? Nous allons le découvrir progressivement dans la suite du texte.
Commençons par définir la notion de rendement[1] quantique de la photosynthèse (Φ). Il s'agit du rapport entre le nombre de photons utilisés pour la photosynthèse et le nombre total de photons absorbés par l'échantillon. Par exemple, si A représente la vitesse d'émission d'oxygène (unité de A : nombre de molécules s-1) par un échantillon absorbant I photons par seconde, alors le rendement quantique peut être calculé à l'aide de l'équation 1
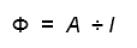
L'unité de Φ est mole O2 (mole photon)-1. Mesurons maintenant la variation du rendement quantique de la photosynthèse en fonction de la densité de photons incidente. Pour ce faire, nous pouvons utiliser le dispositif expérimental présenté à la figure (Figure 23A). L'expérience historique a été réalisée par le Dr EMERSON sur la microalgue verte Chlorella. Un échantillon de microalgues est illuminé par une densité de flux de photons. Les vitesses d'émission de l'oxygène induit par l'illumination est mesurée ainsi que la vitesse d'absorption des photons. L'opération est ensuite répétée autant de fois nécessaire pour construire la courbe représentant la variation du rendement quantique de la photosynthèse en fonction de la vitesse d'absorption des photons (Figure 23B). Dans la pratique, c'est souvent le rendement quantique apparent qui est mesuré car la vitesse d'absorption des photons n'est pas mesurée. Seule la densité du flux de photons est considérée.
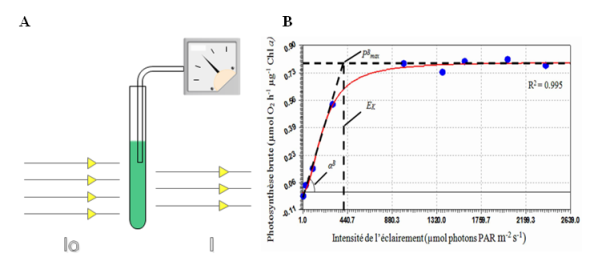
A. Schéma expérimental permettant la mesure du rendement quantique de la photosynthèse. Un échantillon de microalgues est illuminé par une densité de flux de photons. Les vitesses d'émission de l'oxygène induit par l'illumination ainsi que la vitesse d'absorption des photons, sont mesurées.
B. Variation de la vitesse de la photosynthèse brute en fonction de l'intensité de la densité de flux de photons. Les points expérimentaux sont représentés en bleu. La variation de la vitesse de la photosynthèse brute a été mesurée avec des aliquotes de culture de la diatomée Phaeodactylum tricornutum cultivée sous un éclairement de 300 µmoles de photons m-2 s-1. Chaque aliquote a été soumis à un éclairement d'intensité différente. A partir de ces données, plusieurs paramètres caractéristiques peuvent être déduits à savoir αB, PBmax et EK. Le paramètre αB ou rendement quantique apparent a la même dimension que le rendement quantique Φ. Il est apparent car le plus souvent, et c'est le cas ici, la vitesse d'absorption des photons n'est pas mesurée. Seule la densité du flux de photons est considérée. PBmax représente le rendement quantique maximal pouvant être atteint dans les conditions expérimentales dans lesquelles l'expérience a été effectuée. EK correspond à la valeur nominale de la densité du flux de photons pour laquelle la photosynthèse est saturée.
A partir de ces données et de la valeur de Φ, nous pouvons calculer le nombre de photons nécessaires à l'émission d'une molécule d'oxygène. En effet, l'inverse de Φ à cette dimension (Equation 2).
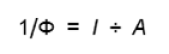
L'unité de Φ-1 est la mole de photons (mole O2)-1. Les valeurs les plus fréquemment trouvées varient entre 8-9. Ce résultat indique que l'émission d'une molécule d'oxygène nécessite l'absorption de 8 à 9 photons. Les travaux menés par Gaffron et Wohl dans les années 1930 ont montré qu'une même molécule de chlorophylle d'une plante illuminée par une lumière de faible intensité ne pouvait absorber un photon qu'une seule fois au cours d'une période de plusieurs minutes (Gaffron et Wohl 1936). Ceci suggère que les molécules de pigments responsables de la capture de la lumière pour la photosynthèse doivent être regroupés. Les expériences réalisées par Emerson et Arnold au cours de la même période ont permis de calculer que l'émission d'une molécule d'oxygène impliquait 2500 molécules de chlorophylles. Mettons en relation ce résultat avec celui de Gaffron et Wohl. Il est aisé de déduire qu'il faut environ 300 molécules de chlorophylle pour capter un photon. Ces groupes de pigments sont regroupés au sein d'une structure appelée l'antenne collectrice de l'énergie lumineuse (en anglais “Light Harvesting Complex” ou LHC) (Figure 24). Ces structures peuvent être isolées des membranes photosynthétiques pour être étudier. Il sort du cadre de cette ressource de détailler la composition de l'antenne collectrice, il suffit de se rendre compte qu'elle contient des pigments associés à des protéines. Il existe deux types d'antenne collectrice de l'énergie lumineuse
l'antenne extramembranaire : se retrouve chez les algues rouges et certaines cyanobactéries en accompagnement d'une antenne intramembranaire de petite taille. L'antenne extramembranaire est désignée par le terme phycobilisome. Les pigments associés à l'antenne extramembranaire dans ce type d'antenne sont des tétrapyrroles ouverts (voir section "Les tétrapyrroles ouverts"). Les pigments sont liés aux protéines par des liaisons covalentes.
l'antenne intramembranaire : se retrouve chez tous les organismes photosynthétiques eucaryotes. Ce type d'antenne contient des molécules de chlorophylle et de caroténoïdes. Elles ne sont pas liées de manière covalente aux protéines. Les dosages montrent que chez les plantes supérieures, l'antenne collectrice contient environ 300 molécules de chlorophylle (a+b) auxquelles s'ajoutent des caroténoïdes.
Le rôle de l'antenne collectrice de l'énergie lumineuse est d'absorber les photons et de les transmettre au centre réactionnel. Nous découvrirons en détail à quoi correspond ces deux entités un peu plus tard. Pour l'instant, nous pouvons nous contenter d'admettre qu'il s'agit d'une structure composée de complexes protéines-pigments dont la fonction est de générer des électrons qui seront injectés dans la chaîne de transporteurs des électrons. D'une façon imagée, il s'agit d'un système ("trap" dans la figure 24) contenant de la chlorophylle a capable de piéger l'énergie de l'antenne. L'énergie est utilisée pour extraire un électron au donneur d'électrons ("D" dans la figure 24) qui est donné à un accepteur d'électrons ("A" dans la figure 24).

Les photons sont capturés par les pigments - des molécules de chlorophylles et de caroténoïdes - de l'antenne collectrice de l'énergie lumineuse. L'énergie associée aux photons est transmise (flèche rouge) à la paire spéciale de chlorophylle (P680 ou P700) du centre réactionnel. L'énergie est utilisée pour extraire un électron d'une de ces molécules de chlorophylle. Celui-ci est donné à un accepteur d'électrons (A) qui s'en trouve réduit. Le vide électronique localisé au niveau de la paire spéciale de chlorophylle est comblé à l'aide d'un électron donné par le donneur (D). Celui-ci s'en trouve oxydé. La flèche rouge représente le cheminement de l'énergie associée au photon absorbé à l'intérieur de l'antenne jusqu'au centre réactionnel. Ce cheminement est aléatoire. Généralement, l'énergie aboutit au niveau du centre réactionnel.
Avant d'aller plus loin dans la découverte du fonctionnement de l'appareil photosynthétique, nous devons comprendre comment l'énergie absorbée se déplace d'un pigment à un autre dans l'antenne collectrice de l'énergie lumineuse et ce jusqu'au centre réactionnel (Figure 24). Le mécanisme s'appelle le transfert d'énergie. Pour commencer, décrivons les acteurs:
il en faut au moins deux à savoir un donneur et un accepteur.Notons le donneur d'énergie, D, et l'accepteur, A (Figure 25A). S'il y a plus de deux participants, l'accepteur Ai peut servir à son tour de donneur (Di) et transférer l'énergie absorbée par D à un accepteur final (Af) (Figure 25B). Voyons comment le transfert d'énergie fonctionne. Si la molécule D se trouve seule en solution, l'excitation de D (donor pigment absorption dans la figure 25C) sera rendue dans l'environnement, par exemple sous forme de fluorescence (donor pigment fluorescence dans la figure 25C). L'introduction dans la solution contenant la molécule D de l'accepteur A va permettre le transfert si
(1) Le spectre d'absorbance de l'accepteur se trouve à plus grandes longueurs d'onde que le spectre d'émission de fluorescence du donneur. Cette condition doit être remplie car A ne sera capable d'absorber que de l'énergie correspondant à l'émission de fluorescence (acceptor pigment absorption dans la figure 25C). la compatibilité entre A et D peut être estimée par la mesure de l'intégrale de recouvrement. Cette surface représente le recouvrement (triangle vert sur la figure 25C) entre les spectres d'émission de fluorescence de D et d'absorbance de A ;
(2) A et D doivent sont très proches et
(3) bien orientés l'un par rapport à l'autre.
Dans ces conditions, l'énergie absorbée par D est transférée à A qui la dissipera. L'efficacité du transfert dépend des trois conditions énoncées ci-dessus.
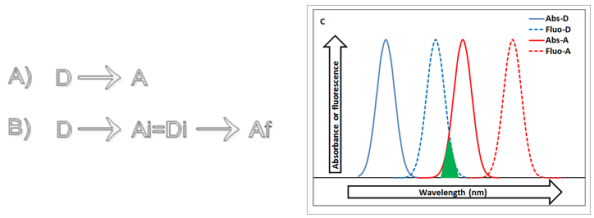
A. Schéma du transfert d'énergie entre deux partenaires, un donneur D et un accepteur A
B. Schéma du transfert d'énergie entre trois partenaires. Dans cette chaîne, l'accepteur Ai peut servir à son tour de donneur (Di) et transférer l'énergie absorbée initialement par D à l'accepteur final (Af)
C. Mécanisme du transfert d'énergie. L'énergie absorbée par la molécule D est transférée à l'accepteur A à condition que son spectre d'absorbance se trouve à plus grandes longueurs d'onde que le spectre d'émission de fluorescence du donneur. La compatibilité entre A et D peut être estimée par la mesure de l'intégrale de recouvrement (en vert).