

8.1 – L'expérience de Spemann et Mangold
L'organisateur de Spemann, situé dans la zone marginale dorsale (ZMD), est induit pendant la période de clivage par le centre de Nieuwkoop, situé dans la région dorsovégétative. Puis, pendant la gastrulation, les mouvements cellulaires redistribuent les territoires embryonnaires, ce qui génère de nouvelles interactions cellulaires dans lesquelles l'organisateur de Spemann est impliqué au premier chef.
L'organisateur de Spemann modifie et oriente la destinée des tissus qui l'entourent. Au niveau du mésoderme latéral, l'organisateur détermine définitivement les dérivés mésodermiques. C'est la dorsalisation. Au niveau de l'ectoderme dorsal, l'organisateur induit le neuroderme, c'est la neuralisation. Quant à l'endoderme, il est vraisemblable que l'organisateur joue un rôle dans sa régionalisation, mais, cela reste à démontrer.
D'un point de vue topographique, l'organisateur de Spemann est inclus dans la zone marginale dorsale. Il se confond avec la lèvre dorsale du blastopore et sa destinée tissulaire est de former principalement la chorde et une partie des somites proximaux (Fig.205).
En réalité, Hans Spemann travaillait avec Hilde Mangold, sa collaboratrice. On devrait donc dire l'organisateur de Spemann et Mangold.
Le rôle particulier de ce 2ème centre décisionnel fut démontré par eux en 1924 après avoir travaillé pendant plus de 10 ans sur ce sujet. Mais, même bien après leur publication, leurs résultats ouvrirent un véritable champ de recherche qui fut couronné en 1935 par un prix Nobel de physiologie et de médecine. La modernité des questions soulevées dans le domaine de la biologie du développement par leurs travaux se prolonge encore, à l'heure actuelle dans les domaines de la biologie cellulaire et moléculaire ainsi que de la génétique.
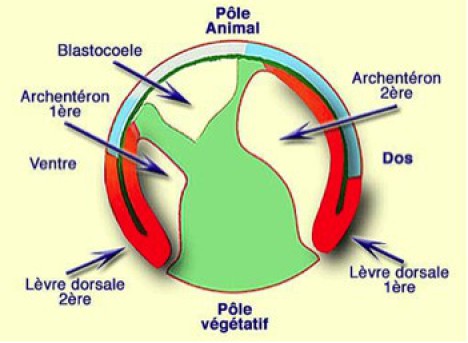
Spemann et Mangold avaient remarqué que, dans une ponte d'amphibien, certains embryons montraient des anomalies du développement corrélées à une perturbation de la gastrulation. Notamment, des embryons exogastrulés ne présentaient pas de face dorsale, ni d'axe antéropostérieur. Ils ont donc supposé que les mouvements de la gastrulation étaient indispensables à la formation de l'axe dorsal et notamment du système nerveux (Fig.206).
Le système nerveux se formant du côté dorsal, leurs soupçons se sont portés sur la lèvre dorsale du blastopore. L'idée leur est donc venue de transplanter la lèvre dorsale du blastopore sur un autre embryon en position ectopique, par exemple dans la région ventrale diamétralement opposée à la lèvre dorsale autochtone.
Le protocole expérimental est le suivant (Fig.207):
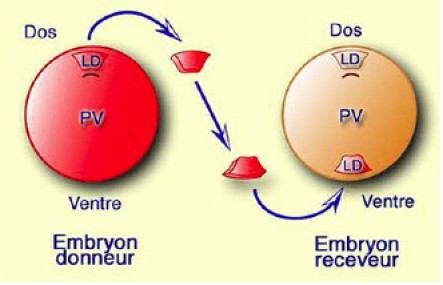
Deux embryons sont choisis au même stade, le début de la gastrulation (encoche blastoporale) . Au préalable, l'un d'eux a été marqué avec du RLDx peu après la fécondation. L'embryon marqué sera le donneur de lèvre dorsale. L'embryon non marqué sera le receveur.
Sur l'embryon receveur, on découpe une fenêtre réceptrice ventrale diamétralement opposée à la lèvre dorsale du blastopore par rapport au pôle végétatif.
Sur l'embryon donneur marqué on prélève la lèvre dorsale du blastopore et on l'implante dans la fenêtre réceptrice de l'embryon receveur. La cicatrisation est complète une heure après l'implantation.
Puis on observe l'évolution de l'embryon (Fig.208). Une coupe sagittale dans l'embryon en cours de gastrulation montre que la gastrulation s'effectue normalement dans la région dorsale de l'embryon receveur. Cependant, dans la région ventrale, un deuxième pôle de gastrulation est initié. La lèvre dorsale implantée s'invagine et recrute les tissus ventraux environnants dans des mouvements de type dorsal.
A cette occasion, un deuxième archentéron se forme du côté ventral. Tout se passe comme si la face ventrale de l'embryon receveur était transformée en face dorsale. Au stade neurula, l'embryon présente sur sa face ventrale, une deuxième plaque neurale opposée à la sienne. Puis, au cours de l'organogenèse un deuxième axe embryonnaire dorsal s'édifie avec des structures troncales et céphaliques plus ou moins élaborées (Fig.209).
Au total, la greffe de la lèvre dorsale du blastopore provoque la formation d'un axe dorsal secondaire. On a réalisé un double axe.
Ce qui importe, c'est que le postulat de départ posé par Spemann et Mangold soit vérifié, car à la suite de la greffe de la lèvre dorsale du blastopore, un système nerveux est bien induit et se développe à partir de l'ectoderme ventral.

En coupe histologique, l'analyse montre que les tissus marqués provenant de la lèvre dorsale du blastopore sont principalement, la chorde, la région proximale des somites, en peu d'endoderme dorsal et quelques cellules du mésenchyme céphalique (Fig.210).
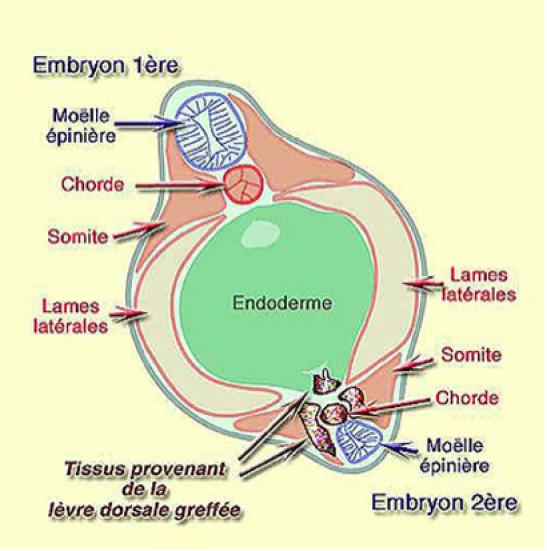
Par contre, le système nerveux n'est pas marqué. Le neuroderme provient donc de l'ectoderme ventral qui a été induit au contact des tissus de la lèvre dorsale, c'est à dire de l'organisateur. Ainsi, l'induction neurale est-elle démontrée dès 1924 par Spemann et Mangold.
Mais il y a plus ! Car si on regarde attentivement la composition des somites de l'embryon induit, on voit très clairement que seule la portion proximale proche de la chorde est marquée et provient donc de la lèvre dorsale greffée.
D'où proviennent les cellules non marquées qui constituent le reste des somites ventraux ?
Il faut nécessairement admettre que les somites de l'embryon ventral aient été induits à partir du mésoderme ventral au contact des tissus de la lèvre dorsale implantée. C'est la dorsalisation du mésoderme ventral. Les cellules, originellement destinées à évoluer en cellules sanguines et en lames latérales haussent leur destinée en mésoderme latérodorsal, c'est à dire les somites. C'est donc bien une dorsalisation.
Comme pour l'induction mésodermique, on peut démontrer que l'induction neurale est régionalisée.
Deux expériences sont possibles :
Soit on greffe la lèvre dorsale du blastopore à différents stades de la gastrulation.
Soit on implante directement dans le blastocoele la lèvre dorsale du blastopore prélevée à différents stades de la gastrulation.
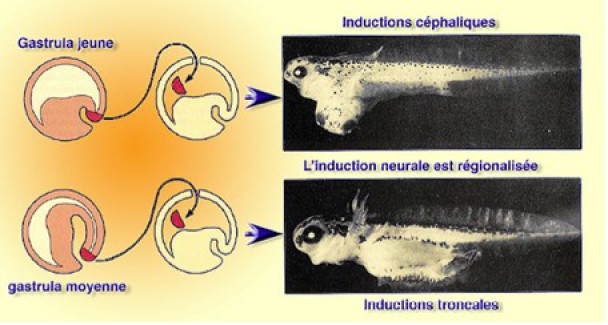
Dans le deuxième cas, on parle d' «einsteck method » (insérer à l'intérieur) (Fig.211). L'explant dans le blastocoele, tombe sur le plancher et est repoussé par les mouvements de la gastrulation vers la région ventrale où il entre en contact avec l'ectoderme. Il se trouve ainsi coincé contre l'ectoderme ventral qu'il induit en neuroderme comme dans le cas de la greffe de Spemann. Dans ces conditions, et compte tenu de la faisabilité de cette technique, on peut pratiquer l'«einsteck method » en grand nombre à partir d'embryons à différents stades.
Dans le premier cas, la lèvre dorsale du blastopore est prélevée au début de la gastrulation.
Dans le deuxième cas, la lèvre dorsale est prélevée vers la fin de la gastrulation (gastrula moyenne).
Les résultats sont les suivants :
Dès l'organogenèse on s'aperçoit que dans le premier cas, on obtient des différenciations principalement céphaliques.
Dans le deuxième cas, les différenciations sont troncopostérieures.
Compte tenu du fait que, pendant la gastrulation, l'invagination de la ZMD internalise les tissus de l'organisateur, en commençant par le mésoderme de tête et en finissant par le mésoderme caudal, on comprend qu'au début de la gastrulation la lèvre dorsale induise des structures céphaliques alors qu'à la fin de la gastrulation, les tissus induits sont caudaux.
L'induction neurale est donc régionalisée.




