Transport à travers la membrane externe
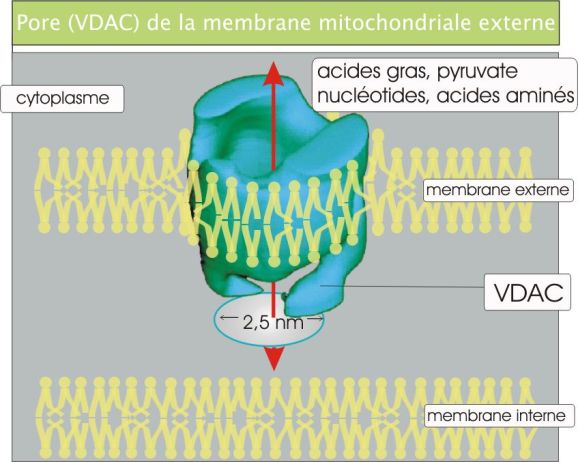
La membrane externe contient des porines, protéines transmembranaires formant de véritables pores. Leur sélectivité et leur spécificité ne sont pas encore bien connues. Le canal anionique VDAC (voltage-dependent anion channel) est considéré comme la voie principale de passage des métabolites à travers la membrane externe. Ce canal est constitué d'une protéine de 31 kDa adoptant une structure de \(\beta-\)tonneau, ressemblant ainsi aux porines de la membrane externe des bactéries (figure 17). Ce pore a un diamètre interne de 2,5 nm qui permettrait donc le libre passage des acides gras, du pyruvate, des acides aminés et de nucléotides.